Splicing process is an adaptative mechanism that allows the cells to increase the flexibility of their transcriptome, therefore playing a key role in tumor evolution and response to treatment (47, 48). Consequently, its dysregulation has emerged as a new key hallmark of cancer development and progression (49). Specifically, our group and others have proven that multiple alternative and aberrant splicing variants emerge in tumor conditions, including PCa (50–52). This altered splicing variant landscape could be a consequence of a marked alteration of the expression levels of spliceosome components and splicing factors in cancer, including PCa (5–8, 53). Interestingly, given the relevance of certain splicing variants in the development, progression, and response to treatment of PCa, splicing regulators play a pivotal role in this pathology (5–9). In this context, the serine/arginine (SR)-rich proteins comprise one of the most relevant families of regulators of the splicing process (10, 11), with many members reported as key factors, such as SRSF1 and SRSF3, have been involved in the progression of several cancer-types, including PCa (5–9). Among the SR family, SRSF6 may represent an important factor based on its involvement in the regulation of several biological processes other than splicing process, such as transcription and protein translation (11, 54, 55).
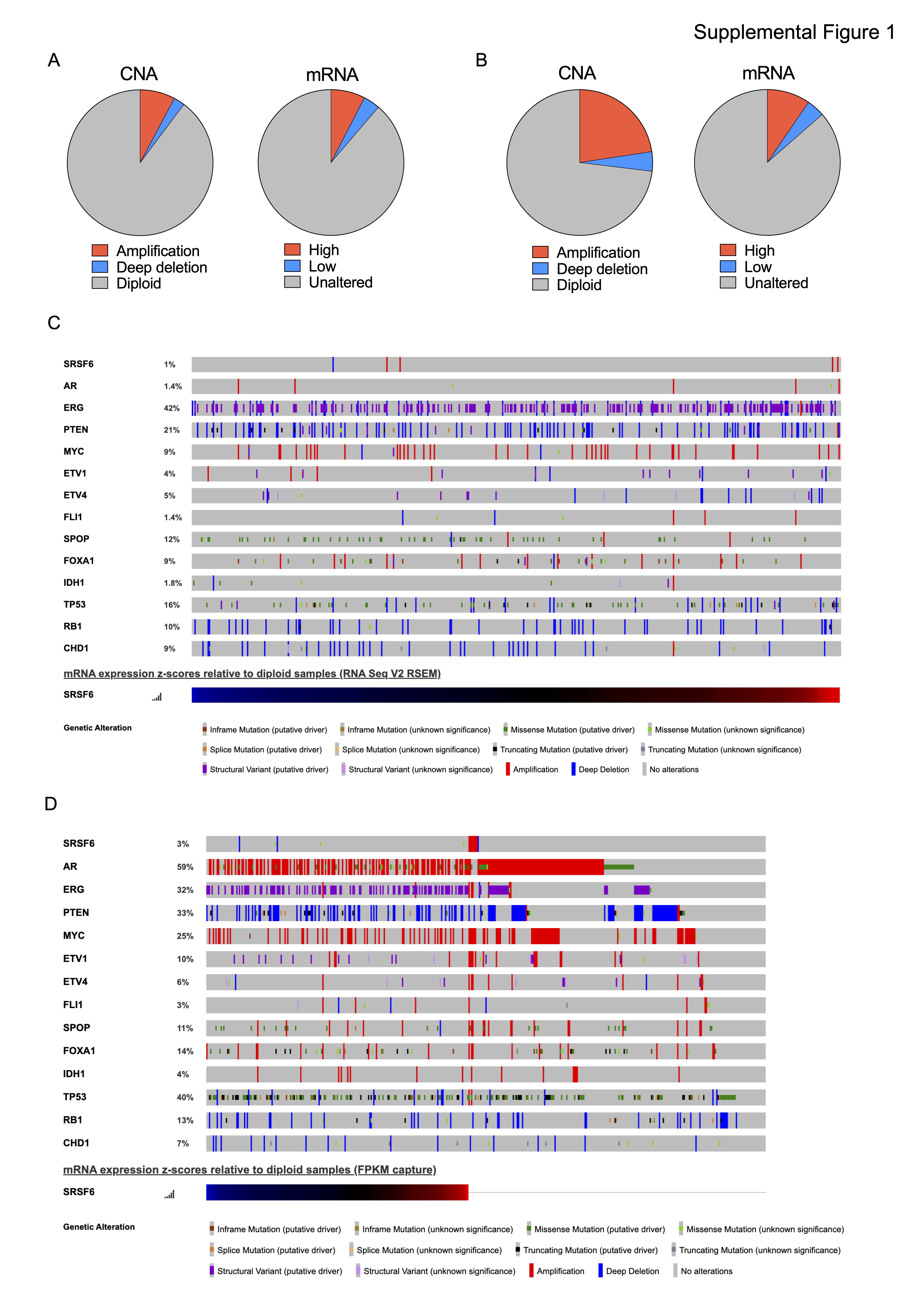
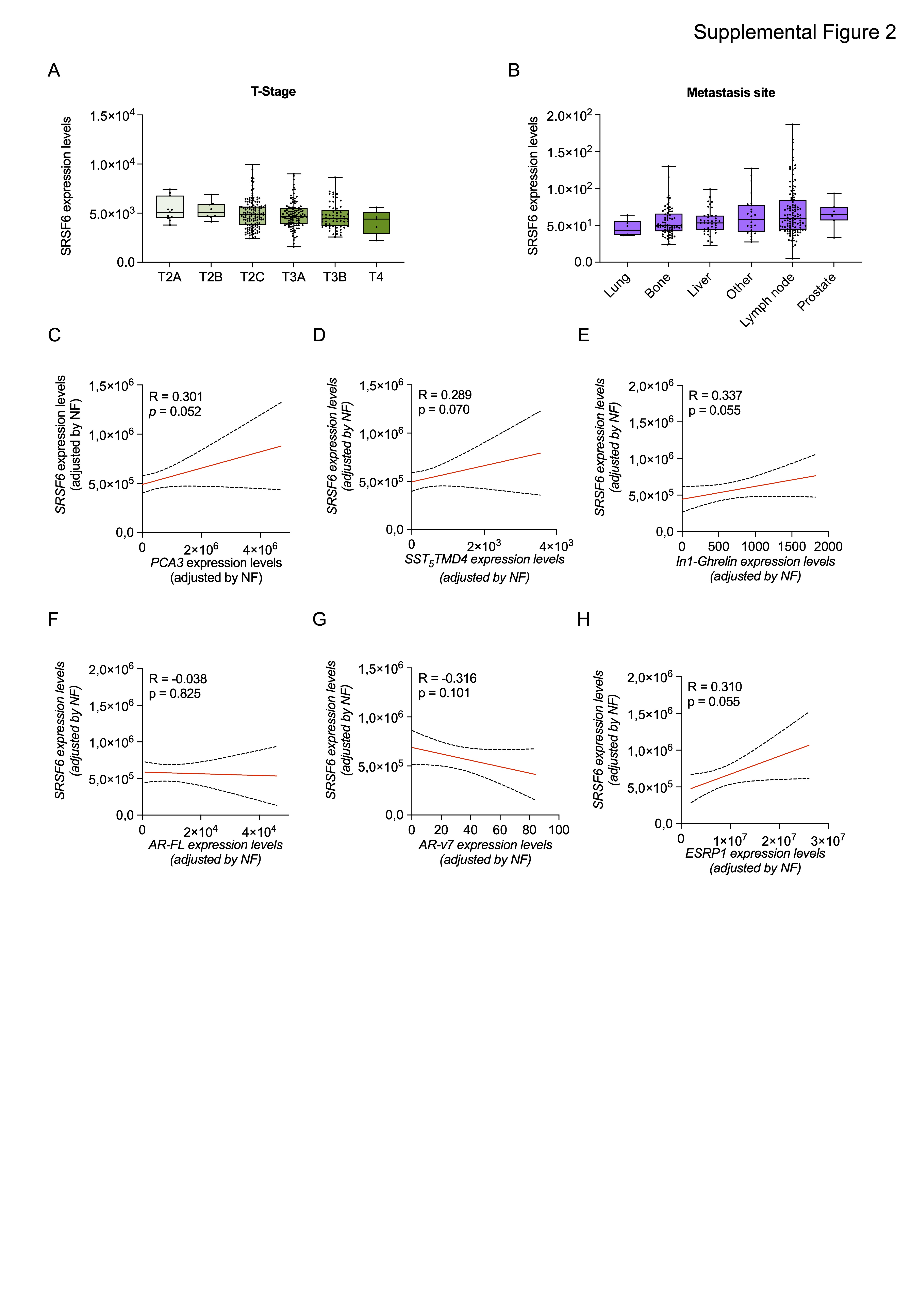
In this study, we analyzed several PCa cohorts to define the levels of SRSF6 and its potential implication in this disease. We demonstrated for the first time that both mRNA and protein levels of SRSF6 levels are higher in PCa tissue as compared to non-tumor prostate or BPH-derived samples, which is in keeping with previous studies showing that SRSF6 is upregulated in certain tumor types, such as melanoma (18), colorectal (19, 20), lung cancer (19) and glioblastoma (56). Consistent with the upregulation at mRNA and protein level, SRSF6 is amplified in a relevant proportion of patients with mCRPC, from the SU2C cohort but also in low-risk early disease from the TCGA cohort, suggesting that SRSF6 amplification-driven upregulation might be also an early event in PCa tumorigenesis. In line with this hypothesis, the amplification of the SRSF6 locus, 20q13, has been proposed as an early event involved in cancer initiation (57). Furthermore, we also found that SRSF6 amplification is more common in highly aggressive disease, mCRPC samples, indicating that SRSF6 upregulation might be also associated with PCa aggressiveness. Consistently, SRSF6 is associated with aggressiveness parameters such as Gleason score, T-stage, perineural infiltration, lymphovascular invasion, and high-volume metastases, in two different cohorts of patients available in our laboratory.
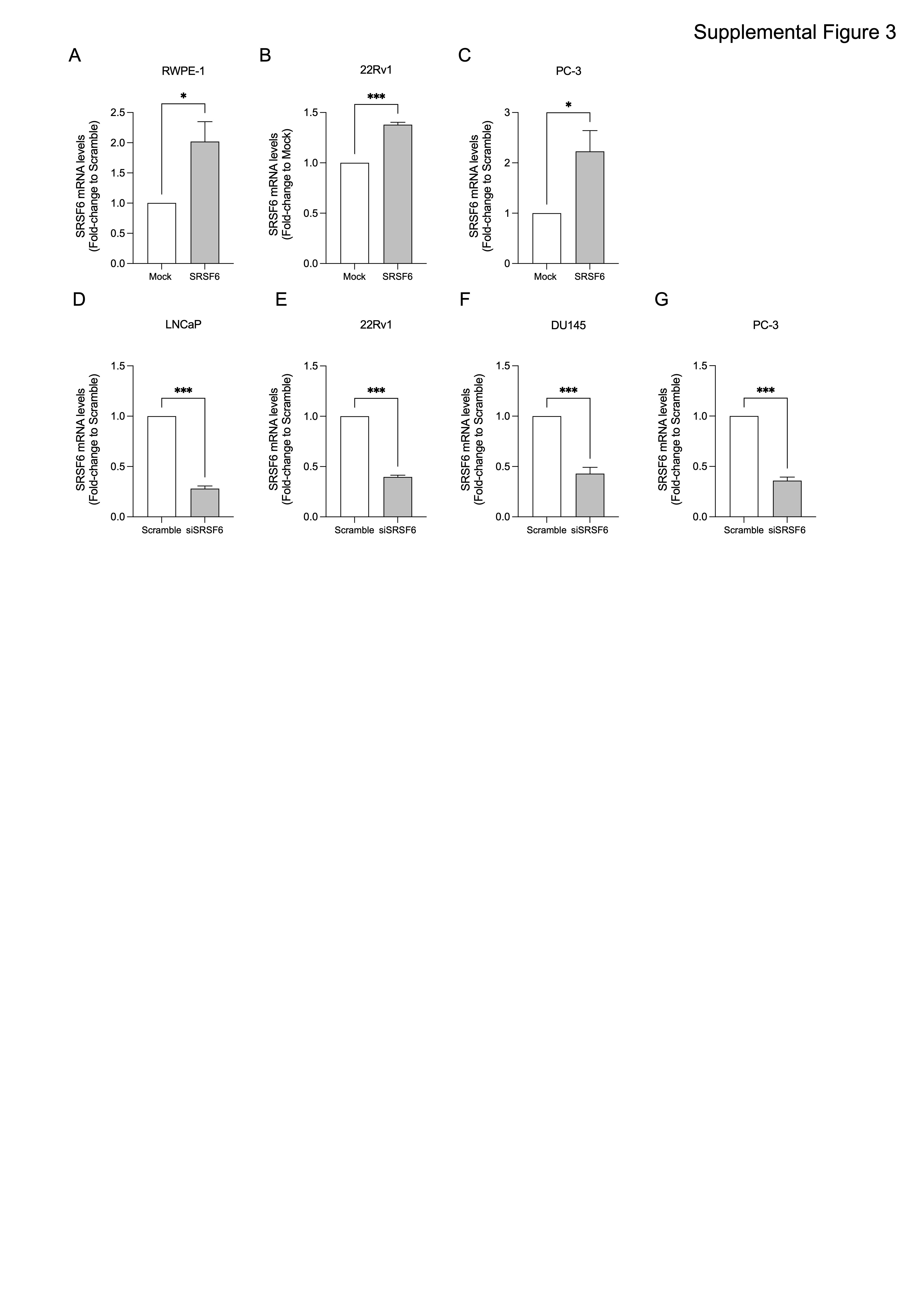
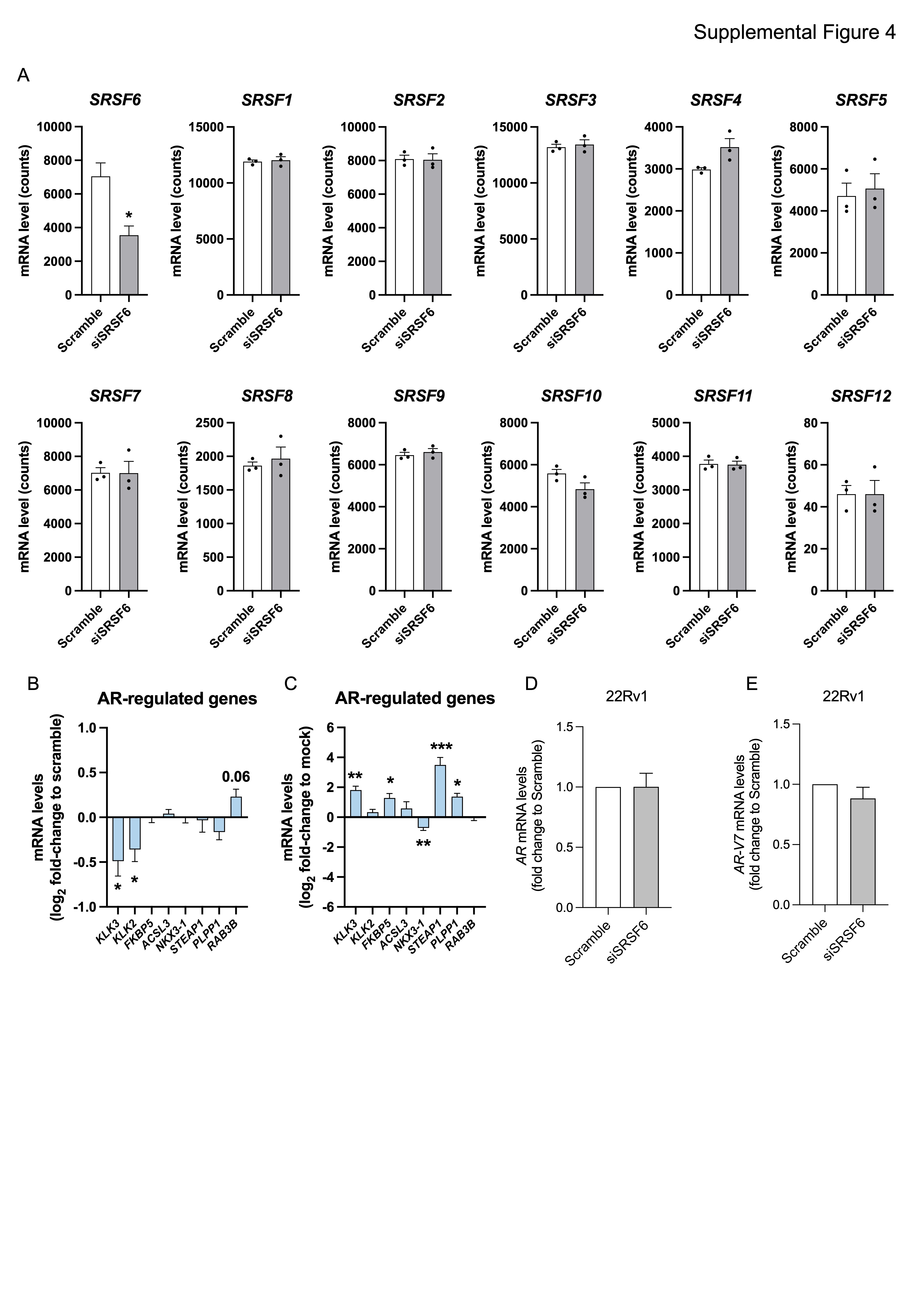
Moreover, we also found that SRSF6 directly modulate PCa cells aggressiveness at the in vivo and in vitro levels. Specifically, the overexpression of SRSF6 increased, while its silencing decreased, functional parameters of tumor aggressiveness including cell proliferation, cell migration, colony and tumorspheres formation. Therefore, these data suggest that SRSF6 plays a relevant pathophysiological role in the progression of PCa, in a similar manner to other SR members, such as SRSF3 (5). Mechanistically, we identified that the “alternative first exon” and “exon skipping” are the most altered splicing event types in response to SRSF6 silencing in PCa cells. Similarly, Wan et al. showed that exon skipping was the most altered splicing type in response to the stable silencing of SRSF6 in colorectal cancer cells (20). These data suggest that some of the mechanisms by which SRSF6 drive tumorigenesis and tumor progression might be shared across different tumor-types, although further experimental evidence is required in this regard. We then cross-compared the pathways altered in response to SRSF6 siRNA in PCa cells and those altered in mCRPC patients with low and high levels of SRSF6 to find potential clinically relevant pathways controlled by SRSF6. As a result, we found that high levels of SRSF6 were associated, among other pathways, with the androgen response hallmark. Consistent with this, SRSF6 mRNA levels correlated positively with a signature of androgen receptor (AR) activity (40) in mCRPC patients. It should be noted that although therapeutic strategies targeting AR still represents the cornerstone of the treatment for patients with advanced PCa, their long-term use is compromised as the tumors eventually develop resistance (2). Therefore, finding novel therapeutic targets that could be exploited to target AR signaling more efficiently and avoid treatment resistance is an urgent unmet clinical need. In this sense, we found that the overexpression of SRSF6 increased, while its silencing reduced, the activity of AR in 22Rv1 cells, a CRPC model resistant to AR inhibition (58). Interestingly, SRSF6 modulation did not result in a change in the expression levels of neither AR nor AR-V7, therefore suggesting that SRSF6 might control AR activity through the modulation of certain regulator(s) of this receptor, such as co-regulators, pioneer factors, etc. In keeping with this, we found that SRSF6 directly control the expression of TOP2B, a topoisomerase required for efficient androgen-mediated gene expression (45), while negatively regulates APPBP2 expression, which promotes cytoplasmatic retention of AR (46). These data indicate that SRSF6 control the expression of key regulators of AR activity and might modulate androgen response by upregulating TOP2B and downregulating APPBP2, suggesting that SRSF6 might represent an exploitable therapeutic target to efficiently block persistent AR signaling in advanced PCa patients. Nevertheless, we also found that SRSF6 levels were associated with additional major tumor-related signaling pathways, other than AR activity. For instance, TP53 and apoptosis pathways (59, 60) were enriched in SRSF6 silenced cells and patients with low SRSF6 expression levels, suggesting that these pathways might be involved in the antitumor effects observed in response to SRSF6 silencing in PCa cells, which may explain the sensitivity of AR negative cell lines (PC-3 and DU145) to SRSF6 depletion. Besides, high SRSF6 levels (in both 22Rv1 cell line and patient samples) were associated with a higher expression of E2F target genes. In this sense, it is widely known that the hyperactivation of the CDK–RB–E2F axis leads to uncontrolled proliferation and is a common feature of highly-aggressive cancer types, including advanced PCa (61). The association between SRSF6 and E2F pathway, in combination with the fact that patients with high SRSF6 levels had worse biochemical free survival in our study, highlights a potential prognostic value for SRSF6 in PCa which could be exploitable clinically.
Taken together, our results unveiled new conceptual and functional avenues in PCa, with potential therapeutic implications, by demonstrating for the first time that SRSF6 is upregulated in PCa which is likely of clinical interest, inasmuch as this alteration directly associates to critical aggressiveness features of PCa. Moreover, we unveil a role of SRSF6 in crucial pathophysiological processes of PCa, such as cell proliferation, colony and tumorspheres formation, and cell migration. These actions are likely mediated through the modulation of key oncogenic pathways in PCa including AR, TP53, E2F, and apoptosis. Therefore, our study provides solid, convincing evidence demonstrating that SRSF6 has a functional role in the pathophysiology of PCa and invites to suggest that SRSF6 may represent an effective prognostic biomarker and exploitable therapeutic target in PCa, offering a clinically relevant opportunity that should be tested for their use in humans.
{kind=link}
{kind=link}
{kind=link}
{kind=link}