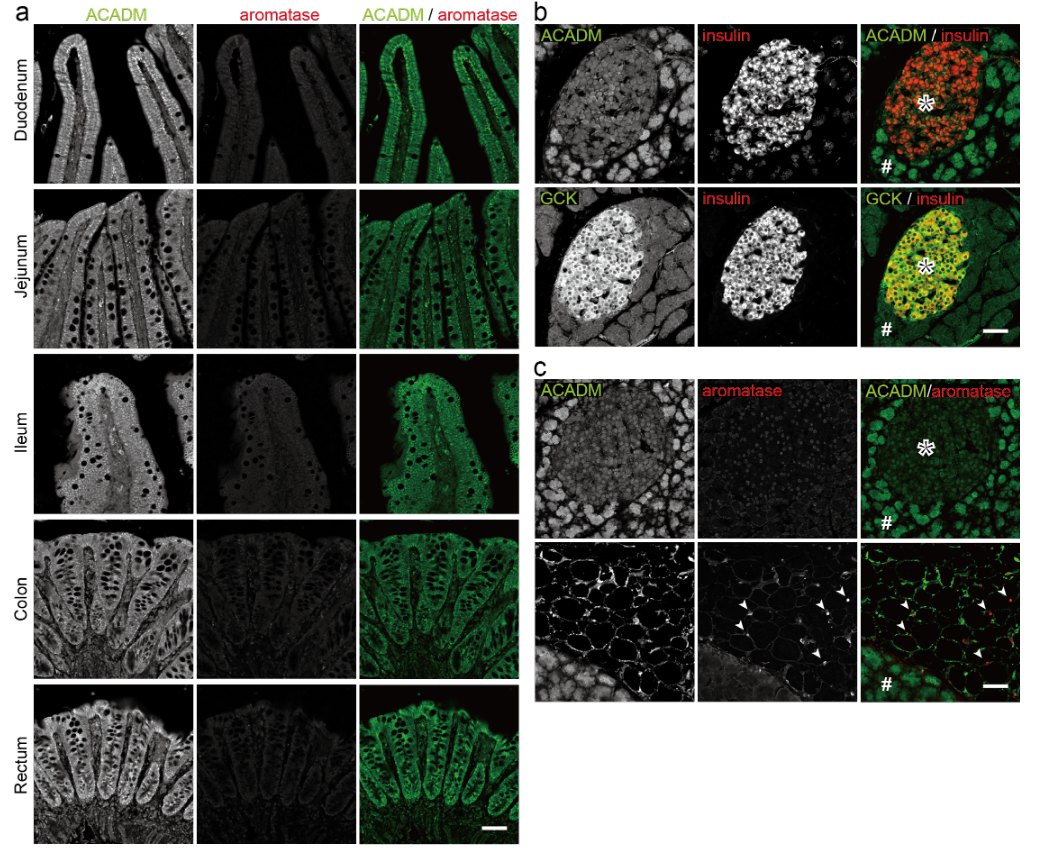
Estrogen is a multi-target and multi-functional hormone beyond just a sex hormone; it controls glucose/lipid homeostasis, bone metabolism, brain function and skeletal growth as well as follicular growth and ovulation7. Aside from the ovaries in females, estrogen is also secreted from adipose tissues, which are greatly increased in males10,11. Our previous studies showed that parietal cells in rat stomachs also express aromatase, together with 17a-hydroxylase and 17b-hydroxysteroid dehydrogenase type III (17b-HSD type III), and convert testosterone (male) or progesterone (female) to estrogen (17b-estradiol [E2])6,12 (Fig. 1a). Aromatase was prominently expressed in the stomach among the tissues upstream of the portal vein (stomach, duodenum, jejunum, ileum, colon and pancreas)6, and the small amounts of aromatase in the rest of the tissues were mainly from the attached adipose tissues (Fig. 1b, c, Extended Data Fig. 1). Parietal cells secrete estrogen (gastric estrogen) into the portal vein, so estrogen levels are higher in the portal vein (ca. x2) than in the artery in male6 and female13 rats, which is lost in gastrectomized (GX) rats6. The role of gastric estrogen and how its secretion is regulated are not widely understood. Here, using male rats, we show that parietal cells act as a sensor for the blood triglyceride levels and gastric estrogen is secreted in response to the blood triglyceride levels. Estrogen suppresses the lipid supply by inhibiting the feeding behavior and the de novo lipogenesis in the liver8,9, so we propose that when blood triglyceride levels rise, parietal cells increase the secretion of estrogen, which lowers the blood triglyceride levels to maintain proper blood triglyceride levels in males (Fig. 3d).
Gastric estrogen directly enters the liver via the portal vein, where estrogen controls glucose/lipid homeostasis9. This secretion is similar to that of insulin, which is secreted from pancreatic b-cells, enters the portal vein, and directly acts on the liver to keep appropriate blood glucose levels14,15. b-cells, which sense the blood glucose levels, use glucose, not fatty acids, as an energy source16 (Extended Data Fig. 1b), and parietal cells require energy (NADPH) in the production of estrogen (3 x NADPH from testosterone in male, 6 x NADPH from progesterone in female, Fig. 1a). We therefore focused on the energy sources of parietal cells to obtain clues on how the production of gastric estrogen is regulated. Blood estrogen levels change in the menstrual cycle in females, so in this study we used male rats to exclude the effect of estrogen from the ovaries. Triglyceride (fatty acids) and glucose are major sources of energy for mammals. We examined the expression of the enzymes used in the generation of energy from fatty acids (acyl-CoA dehydrogenase medium chain [ACADM], an enzyme involved in b-oxidation) and glucose (glucokinase [GCK], a glycolytic enzyme) in the gastric mucosa (Figs. 1c, d). We identified the estrogen-producing cells and parietal cells using the antibodies against aromatase and H+/K+ ATPase (ATP4B), respectively6. Aromatase-positive cells expressed high levels of ACADM, but little if any GCK. Conversely, aromatase-negative cells showed strong GCK but weak ACADM expressions (Fig. 1c). ATP4B-positive parietal cells showed the same result as the aromatase-positive cells (Fig. 1d). We therefore conclude that parietal cells exclusively produce estrogen in the stomach and predominantly use fatty acids as an energy source.
Parietal cells secrete estrogen in an energy-dependent manner6,12 (Fig. 1a), and they predominantly use fatty acids as an energy source (Fig 1c and d). Moreover, estrogen from parietal cells directly enters the liver via the portal vein where lipogenesis occurs when blood triglyceride levels are low17, and it suppresses the increase of blood triglyceride levels by inhibiting the hepatic de novo lipogenesis9 and the feeding behavior8. We therefore hypothesized that parietal cells increase the secretion of estrogen as blood triglyceride levels rise, which suppresses the supply of lipid to maintain appropriate blood triglyceride levels.
To test this hypothesis, we designed a series of experiments. First, we investigated the relationship between the blood triglyceride and estrogen levels (Fig. 2). Triglyceride (olive oil, 2.5 ml/kg body weight) or control water was orally administered to rats and we measured their triglyceride and estrogen (E2) levels in the tail venous blood before (0 h) and at 1, 2, 3, 4, 5 h after the administration (Fig. 2a). Blood triglyceride levels (basal levels: ~130 mg/dL) increased, peaked at 2 h post-administration (~300 mg/dL), and then declined to the basal levels (5 h) as reported previously18. Blood E2 levels (basal levels: ~50 pg/ml) also increased, peaked at 2 h after the administration (~80 pg/ml), and then returned to the basal levels (5 h). Control rats did not show apparent changes in their blood triglyceride or in E2 levels. We then analyzed the relationship between the blood triglyceride and E2 levels at 2 h after the administration; there was a positive correlation between them (Fig. 2b). We further examined the effect of blood glucose on the blood estrogen levels. Glucose administration is known to decrease blood triglyceride levels via insulin-mediated inhibition of lipolysis in adipose tissues19,20, so we also monitored the blood triglyceride levels. Rats were orally administered glucose (2 g/kg body weight) or control water, and we measured their tail venous blood glucose, E2, and triglyceride levels before (0 h) and at 0.5, 1, 1.5, 2, 3, 4, 5 h after administration (Fig. 2c). Blood glucose levels (basal levels: ~60 mg/dL) increased, peaked at 1 h after the administration (~170 mg/dL), and then declined toward the basal levels21. Blood E2 and triglyceride levels did not increase, however, they actually decreased when blood glucose levels were high. Blood estrogen levels are indicated to increase as blood triglyceride levels, but not glucose levels, rise.
Next, we investigated the effect of blood triglyceride levels on the production of estrogen in parietal cells. Parietal cells exclusively produce estrogen in the stomach and the elevated estrogen level in the portal vein compared with the artery is cancelled in GX rats6. We therefore measured the E2 levels in the stomach tissues with the triglyceride levels in the tail venous blood at 2 h after the administration of olive oil or control water (Fig 3a). Rats administered olive oil showed elevated blood triglyceride (~280 mg/dL) and stomach E2 (~3,500 pg/g tissue) levels compared with the controls (blood triglyceride: ~110 mg/dl, stomach E2: ~1,900 pg/g tissue), and there was correlation between the blood triglyceride and stomach E2 levels. Parietal cells in the stomach were shown to produce estrogen in response to the blood triglyceride levels.
Finally, using GX rats, we investigated the involvement of the stomach in the regulation of blood estrogen levels by blood triglyceride levels (Fig. 3b, c). GX rats showed weaker increase of the blood triglyceride levels than normal rats when administered olive oil (data not shown), so we used 5 ml/kg body weight of olive oil in the studies using GX rats. Five days after total gastrectomy, GX rats were administered olive oil or control water. We measured triglyceride and E2 levels in the tail venous blood before (0 h) and at 1, 2, 3, 4, 5 h after the administration (Fig. 3b). Blood triglyceride levels of GX rats (basal levels: ~90 mg/dL) increased, peaked at 2 h post-administration (~210 mg/dL), and then declined to the basal levels (5 h); but, blood E2 levels did not show apparent changes (~60 pg/ml). We also analyzed the relationship between the blood triglyceride and E2 levels at 2 h after the administration, but we found no correlation between them (Fig. 3c). The stomach is indicated to be the organ responsible for controlling the blood estrogen levels in response to the blood triglyceride levels.
In conclusion, we showed that parietal cells in the stomach predominantly produce estrogen among the tissues upstream of the portal vein. Production of estrogen requires energy, and parietal cells predominantly use triglyceride as the energy source. Blood estrogen levels increase as blood triglyceride levels rise, but not as glucose levels rise. Levels of estrogen in the stomach tissues increase as blood triglyceride levels rise, and in GX rats, increases of blood estrogen levels dependent on blood triglyceride are cancelled.
To maintain proper lipid conditions, hypothalamic NPY neurons collect the body lipid status to control hepatic lipogenesis and feeding behavior1. Ghrelin from the stomach, leptin from adipose tissues and vagus afferent nerves from the intestines and the liver report the levels of fasting, body fat and ingested lipids, respectively, and play a part in activation (ghrelin) or inhibition (leptin and vagus afferent nerves) of the NPY neurons2,4,5. Little is known, however, about the hormones or nerve fibers that inform the blood lipid levels. On the other hand, estrogen inhibits the hepatic de novo lipogenesis and the subsequent secretion of VLDL directly9,22,23 or by suppressing the NPY neurons8,24,25. Little is known, however, about what information blood estrogen levels represent in terms of the lipid homeostasis. Among the organs upstream of the portal vein, the stomach and the pancreas do not take up nutrients and are suitable for sensing and directly informing the liver of the levels of “circulating” nutrients. Indeed, the pancreas secretes insulin or glucagon depending on the circulating blood glucose levels to control the blood glucose levels primarily by acting on the liver. We therefore collectively conclude that the stomach, a digestive organ, also acts as a blood triglyceride sensor organ, and parietal cells in the stomach secrete estrogen according to the blood triglyceride levels (Fig. 3d). When blood triglyceride levels rise, parietal cells increase the secretion of estrogen into the portal vein, which in turn lowers the blood triglyceride levels by inhibiting the hepatic de novo lipogenesis and feeding behavior. Conversely, when blood triglyceride levels lower, parietal cells decrease the secretion of estrogen to supply more lipid to the circulation.
This is the first demonstration of a mechanism by which gastric parietal cells sense the blood lipid levels and secrete estrogen to keep proper blood triglyceride levels by controlling the hepatic de novo lipogenesis and feeding behavior. Our model is consistent with previous reports on the daily rhythms of hepatic lipogenesis, blood triglyceride levels, and blood and gastric estrogen levels. Hepatic lipogenesis is regulated by daily feeding rhythm and rises when blood triglyceride levels are low17. Blood estrogen levels increase at night when rats eat actively, which is regulated by the secretion of gastric estrogen26.
Adipocytes dominantly use fatty acids as the energy source like parietal cells, and express high levels of ACADM (Extended Data Fig. 1c) and low levels of GCK (data not shown). Blood triglyceride-dependent changes of blood estrogen levels are regulated by the stomach, but not by other tissues, such as adipose tissues. Adipocytes are filled with triglyceride, but parietal cells are not. Intracellular triglyceride levels are thus more susceptible to the extracellular (blood) triglyceride levels in parietal cells than in adipocytes. We therefore think parietal cells, not adipocytes, are suitable for the real-time monitoring of blood triglyceride levels.
As for gastric estrogen in females, parietal cells of female rats also secrete estrogen and increase the estrogen levels in the portal vein13; but estrogen and progesterone, a precursor for gastric estrogen in females, are mainly secreted from the ovaries. Moreover, the blood levels of estrogen and progesterone change dramatically with the menstrual cycle. The roles and regulation of gastric estrogen in females require further careful investigation.
{kind=link}