3.1 Expression profile of the splicing machinery is altered in pulmonary carcinoids, enables to discriminate tumor vs. non-tumor tissue, and unveils new molecular links with clinical features
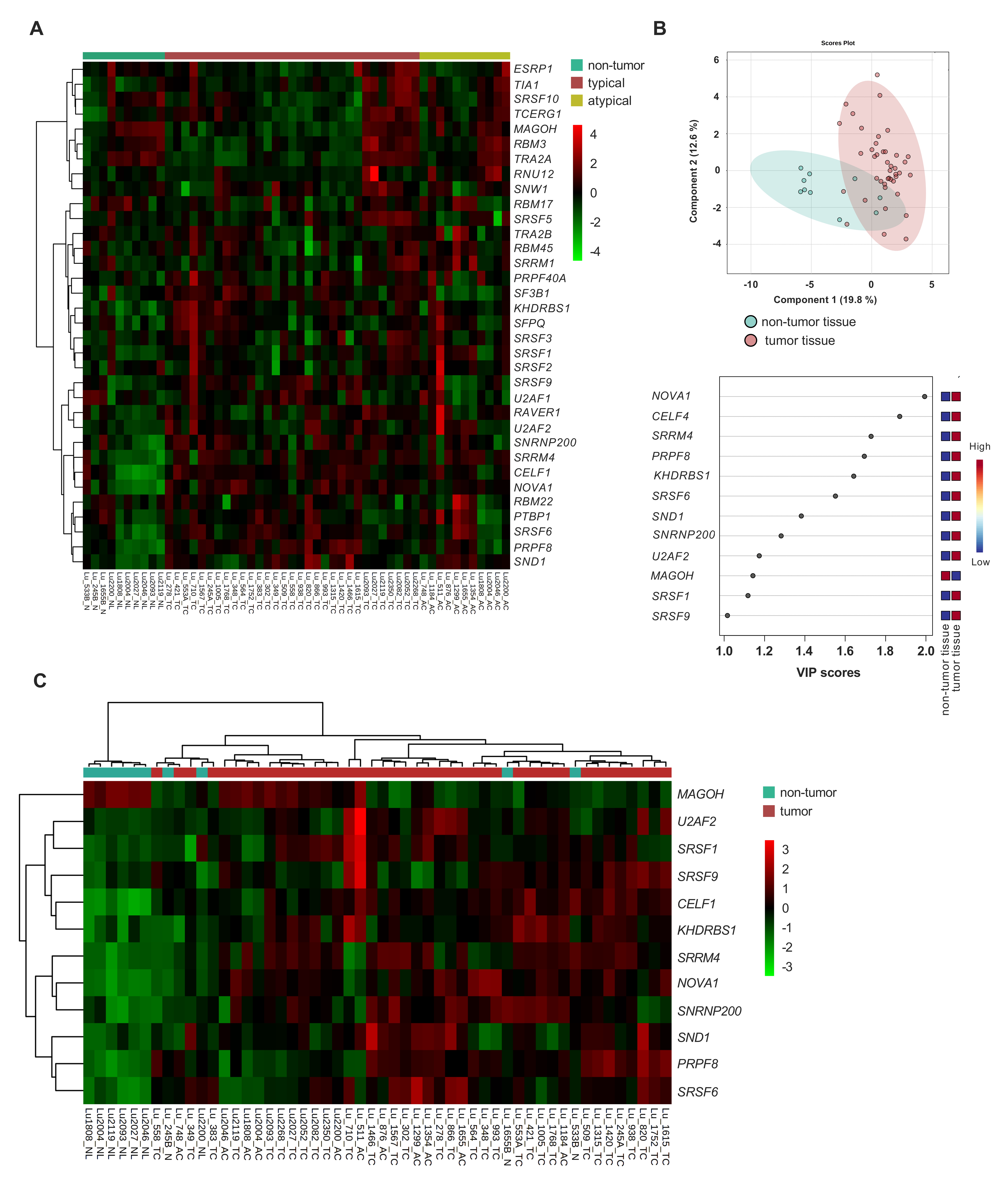
The expression of 10 of the 43 splicing machinery components evaluated (23.3%) was altered in pulmonary carcinoid tissue when compared to their respective non-tumor adjacent tissue (Wilcoxon test, p < 0.05; Fig. 1A). Specifically, the splicing machinery components KHDRBS1, NOVA1, PRPF8, SNW1, SRSF1, SRSF10 and SRSF9 were overexpressed in tumor tissue. Moreover, in the core of the spliceosome machinery, the snRNAs RNU4-1, of the major spliceosome, and RNU12 and RNU4ATAC, of the minor spliceosome, were also overexpressed in tumor tissue. No overt differences were observed between the two carcinoid subtypes (Fig. 1B, see Histology in Additional file 2: Fig. S2). To examine these results in more detail, we analyzed an external validation cohort (GSE108055, Additional file 1: Fig. S1A) (39). The dataset explored in this case derives from a mRNA expression microarray, which contains nearly 80% of the genes evaluated in our microfluidic array (34 out of 43 genes), mostly because snRNA (which lack a poly-A tail) were not targeted by this technique. Interestingly, 16 of the 34 components examined (47.1%) were altered and, in line with our discovery cohort, KHDRBS1, NOVA1, PRPF8, SNW1, SRSF1, and SRSF9 were also overexpressed in tumor tissue in this external cohort (Fig. 1C). At this point, to analyze these results with a more objective perspective, we should introduce the caveat that the wide diversity of cell types in the tumor surrounding tissue, together with the low proportion of neuroendocrine cells in bronchial tissue (40) is admittedly a general limitation in the study of these tumors, as it hinders a balanced comparison between the tumor tissue and the adjacent non-tumor component of the tissue. Hence, we routinely consider the neighboring non-tumor tissue more as a reference tissue for comparisons than a bona fide control tissue. Notwithstanding this, the caveat does not preclude comparing both tissues, and, therefore, we applied a customized biocomputational and statistical approach developed for this purpose (41). Specifically, partial least squares discriminant analysis (PLSDA) of the expression data revealed that splicing-related genes were good discriminators of tumor vs non-tumor tissue. Moreover, the Variable Importance in Projection (VIP) Scores allowed to quantify the importance of each splicing-related gene to the discriminant model (Fig. 2A). The application of the same type of analysis to the external validation cohort resulted in a highly similar outcome, in that the expression levels of the splicing-related genes clearly discriminated tumor from non-tumoral tissue and both VIP Scores models displayed a substantial overlap with 5 shared genes (Additional file 1: Fig. S1B). In line with these observations, non-supervised hierarchical clustering using the top 10 discriminant genes according to VIP Scores unveiled two major clusters that were respectively enriched (Fisher’s exact test p = 0.004) in non-tumor and tumor samples (Fig. 2B, Additional file 1: Fig. S1C).
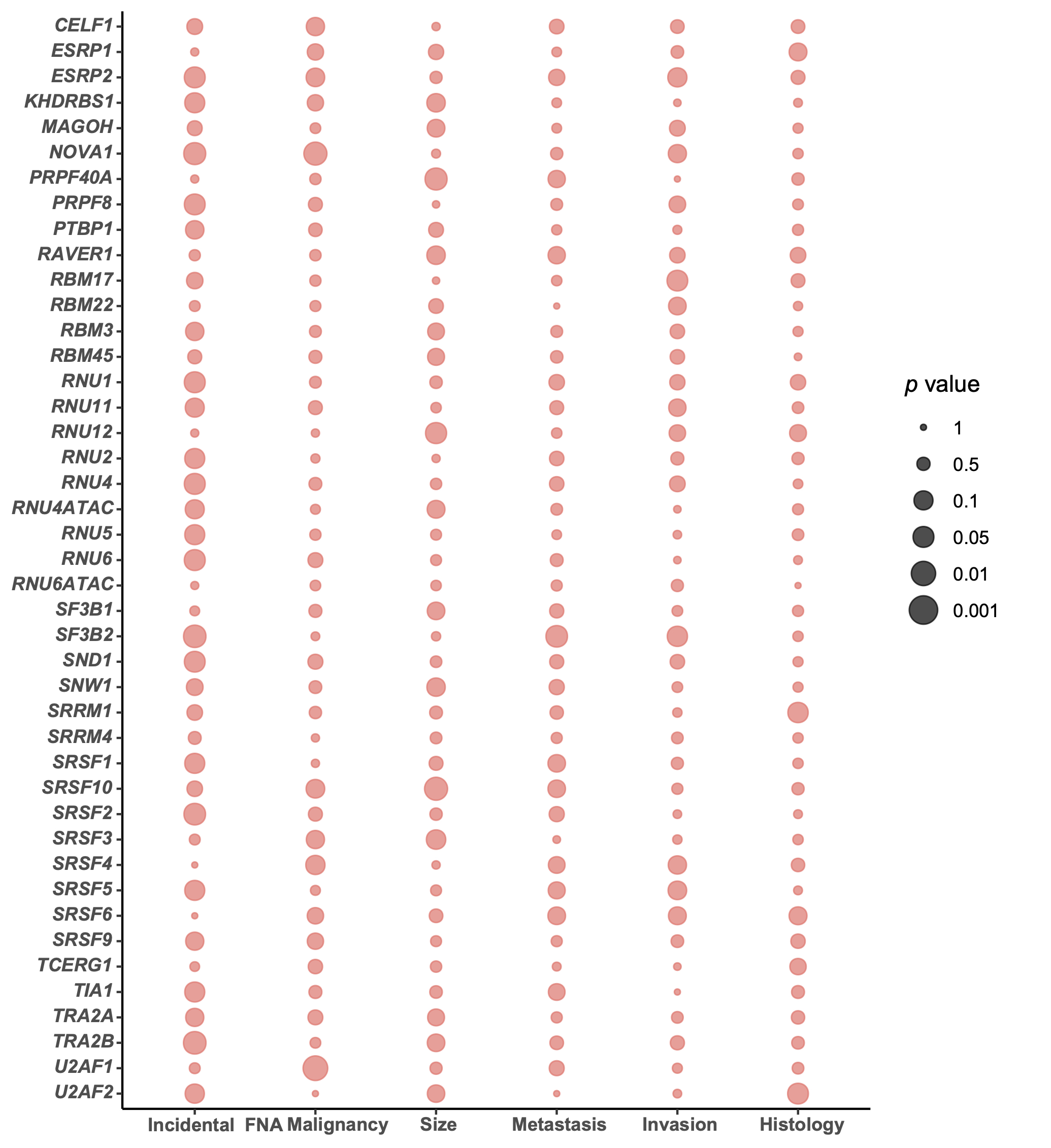
Based on PLSDA and clustering analysis, we selected the top four dysregulated components of the splicing machinery displaying the best discriminating capacity to further explore their role in pulmonary carcinoids, namely: NOVA1, PRPF8, SRSF1 and SRSF9. Of note, these genes were also among the best discriminators of the PLSDA analysis in the validation cohort. Simultaneously, a global screening of the potential associations between the expression levels of each of the splicing factors measured with the most relevant clinical parameters of patients provided an informative snapshot (Additional file 2: Fig. S2), which allowed us to select another interesting component of the splicing machinery, SRSF10, that was also overexpressed in tumor tissue. As illustrated in Fig. 3, these five genes showed similar association profile between their increased expression and incidental diagnosis, reaching statistical significance for NOVA1, PRPF8 and SRSF9. In addition, NOVA1 expression levels were lower when positive malignancy was confirmed after fine needle aspiration, SRSF9 expression was also lower in metastatic disease. Moreover, SRSF10 expression was negatively associated to tumor diameter.
3.2 Protein levels of selected splicing factors unveil heterogeneous distribution in tumor tissue
The presence of the selected splicing factors in carcinoids was further examined by IHC analysis, which confirmed that the protein of three splicing factors, NOVA1, PRPF8 and SRSF10 was detectable in tissue samples. In particular, NOVA1 exhibited a moderate focal cytoplasmic staining and intense but heterogeneous nuclear staining in tumor tissue (Fig. 4A), while, in the adjacent non-tumor tissue, composed of connective tissue and seromucous glands, an almost complete absence of staining was observed. In the case of SRSF10, the tumor tissue showed a mild staining at the cytoplasmic level that contrasted with an intense and uniform staining at the nuclear level, whereas adjacent non-tumor tissue showed very weak staining in the cytoplasm and weak and diffuse staining in the nuclear compartment (Fig. 4B). Likewise, IHC for PRPF8 revealed a moderate staining in the cytoplasmic compartment accompanied by intense staining at the nuclear level in the tumor component of the sample, similar to that described for SRSF10; in contrast, the adjacent non-tumor tissue showed weakly stained cytoplasm and nuclei lacking staining (Fig. 4C). Thus, in line with the RNA expression data, the IHC analysis revealed an overexpression of the three splicing factors NOVA1, PRPF8 and SRSF10. Conversely, application of a similar approach using various methods and antibodies did not reveal consistent differences in the signal abundance and intensity for SRSF1 and SRSF9 in tumor vs. non-tumor tissue.
3.3 NOVA1, PRPF8 and SRSF10 have distinct molecular profiles associated to their expression
To explore in more detail the potential role of NOVA1, PRPF8 and SRSF10 in pulmonary carcinoids, we analyzed a publicly available RNA-seq dataset (EGAD00010001719) from 18 atypical carcinoids. Gene set enrichment analysis (GSEA) performed according to Hallmarks gene sets revealed that the expression of each splicing factor distinctly correlated to a discrete number of hallmarks (Fig. 5A). Thus, whereas NOVA1 was negatively correlated with genes belonging to unfolded protein response, MYC targets, MTORC1 signaling, E2F targets, and G2M checkpoint, the expression of PRPF8 was negatively associated to androgen response, genes downregulated by UV response, Hedgehog signaling, mitotic spindle, TGF beta signaling and G2M checkpoint. In marked contrast, SRSF10 expression was positively correlated to genes that belong to mitotic spindle, heme metabolism, G2M checkpoint, androgen response and Hedgehog signaling. Interestingly, some of the altered pathways, particularly G2M checkpoint, were shared across the three splicing factors.
Inasmuch as the primary known role for NOVA1, PRPF8 and SRSF10 is their function as splicing factors, we aimed at examining their putative relationship with the alternative splicing profile in carcinoid cells. To this end, we calculated the Percent Spliced In (PSI) of alternative splicing events in every tumor sample of the RNA-seq. Samples were classified according to the expression of each splicing factor into high and low expressing samples, and differences in alternative splicing were calculated between both groups. This approach allowed us to assess the potential association between the expression levels of each splicing factor and the pattern of alternative splicing inside the tumor, which could bear functional implications. Interestingly, results unveiled very distinct association patterns for each of the studied factors. Specifically, whereas NOVA1 displayed a reduced set of 35 significantly altered alternative splicing events associated to its low/high expression level (Fig. 5B), the expression of PRPF8 was associated to 2905 significant events (Fig. 5C), and that of SRSF10 to 95 events (Fig. 5D). Differences among splicing factors are not related only to the number but also to the distinct patterns of alternative splicing associated to each of them. Thus, as illustrated in Fig. 5E, whereas NOVA1 was associated to less intron retaining, and more alternative first exon events, PRPF8 displayed an increase of skipping exon events and a clear reduction of first and last exon events, and SRSF10 associated events were enriched in 5’ and 3’ alternative splice sites to the detriment of alternative first exon events.
3.4 Targeting splicing factors in vitro elicits antitumoral effects in lung carcinoid cell models
Having shown the alternative splicing-related features associated to each splicing factor, we next aimed to interrogate the possible functional role played by these factors in pulmonary carcinoids. To this end, since their expression was augmented in tumor tissue, we performed silencing experiments of NOVA1, PRPF8 and SRSF10 in UMC-11 and NCI-H727 cells, two distinct broadly used pulmonary carcinoid cell models (Fig. 6). We first found that, despite their varied levels of expression under basal culture conditions, the silencing of the three factors was comparably effective in each cell line, being overall more pronounced in NCI-H727 with respect to UMC-11 cells (Fig. 6A, 6B). Silencing NOVA1 and SRSF10 decreased NCI-H727 cell proliferation at 72 h and at 48, 72 and 96 h, respectively, when compared to scrambled-transfected cells. However, no effects on cell proliferation were detected in UMC-11 cell line (Fig. 6C). Meanwhile, silencing PRPF8 showed a marked decrease on cell proliferation in both cell lines after 48 h of expression inhibition. Moreover, NOVA1, PRPF8 and SRSF10 silencing also decreased colony formation ability of both UMC-11 and NCI-H727 cell lines, being NOVA1 silencing the one that exerted the highest effect on UMC-11 cells and PRPF8 silencing in NCI-H727 cells (Fig. 6D).
{kind=link}
{kind=link}