Chicken manure (CM) is a major by-product of the poultry industry, consisting of a mixture of chicken faeces and bedding materials (generally sawdust, rice husks and wheat straw) (S.G. Wiedemann, 2015). With the increase in the poultry breeding industry, large amounts of CM biomass are produced annually with an estimated of 21,000 Mt (Jurgutis et al., 2020). This waste product can be used in farmlands as an organic fertiliser. However, a pre-treatment stage is required before its application to the soil, because of the negative impacts on the environment (greenhouse gas emissions, eutrophication and odour arising from say H2S emission ) associated with the direct application of CM to the soil (Chen et al., 2021). Alternatively, CM can be used as a feedstock in anaerobic digestion (AD) to produce biogas which can be used as a fuel source to generate electricity. It has been reported in a study performed in the Netherlands that one tonne of organic waste can produce 222.3 KWh of electricity (Gebrezgabher et al., 2010). Currently, only 30–40% of CM is used for the production of biogas from AD in Asia and Europe (Jurgutis et al., 2020).
The use of livestock manure as a substrate/feedstock in AD is challenging due to a number of constraints, including the accumulation of ammonia (NH3) during the digestion process, the high pH of digestate, and the low C:N ratio of manures (Ngo, Shahsavari, et al., 2022). Among all livestock manures, the AD of CM is considered the most challenging as it has the highest nitrogen (N) content (Forster-Carneiro et al., 2008). The high percentage of N results in the build-up of NH3, which leads to the inhibition of AD and the production of biogas. In fact, the hydrolysis of uric acid and undigested proteins from CM, which are considered the main sources of N in this manure, results in the production of large amounts of amino acids (Molaey et al., 2018). During acidogenesis, these high levels of amino acids release significant concentrations of N which results in a build-up and ultimate accumulation of NH3 (Shi et al., 2017).
Free ammonia nitrogen (FAN/ NH3) and ammonium ions (NH4+) are the two forms of total ammonia nitrogen (TAN) present in aqueous environments (Jiang et al., 2019) and believed to be the main inhibitors of biomethane production during AD (Jiang et al., 2019; Ngo, Shahsavari, et al., 2022). Bujoczek et al. (2000), found that at NH3 concentration above 250 mg L-1, inhibition can occur and methane production will decrease sharply. Ammonia is considered toxic due to its physiochemical properties, such as its high solubility in lipids and its uncharged nature, which allows its diffusion across bacterial cell membranes (Müller et al., 2006). During AD, NH3 can diffuse readily through the cell membranes of methanogens, which helps to maintain the equilibrium of NH3 between the cytosol of bacterial cells and their external environment (Jiang et al., 2019). Within bacterial cells, NH3 acquires a proton by reacting with H+ to form NH4+. In contrast, the diffusion of NH4+ through the cell membranes is not easy as it depends on 2 factors: the intracellular concentration of H+, which determines the pH of the cell, and the extracellular concentration of NH3 (Ngo, Shahsavari, et al., 2022). Consequently, a high concentration of NH3 in the digesters would deter the diffusion of NH4+ which results in its accumulation within the methanogens thus causing a pH imbalance. Furthermore, to balance the proton concentration in the cell, methanogens require more energy to pump K+ out of the cells, which results in the depletion of intracellular K+ (Puig-Castellví et al., 2020). Additionally, the high concentration of NH3 may alter the functioning of K+ pumps, which results in the rapid accumulation of intracellular NH4+ and ultimately a decrease in intracellular pH causing cytotoxicity (Jiang et al., 2019).
Several methods are currently applied to mitigate NH3 inhibition during the AD of CM such as microflora acclimation, trace elements supplementation such as Fe2+ and Ni2+, bentonite addition, C:N ratio adjustment and air stripping (Bi et al., 2019; Chen et al., 2008; Fakkaew & Polprasert, 2021; Ma et al., 2018; Rajagopal et al., 2013). Recently, the use of biochar as additive in AD systems has attracted great interest due to its benefits to the AD process. Biochar is a pyrolytic material, rich in carbon and derived from organic biomass (Lehmann et al., 2006). Several structural and physiochemical properties enable biochar to mitigate NH3 inhibition, including richness in surface functional groups, cation exchange capacity (CEC), and high adsorption and porosity (Chen et al., 2021; J. Ma et al., 2021). Biochar has surface functional groups such as carboxyl (COO-) and hydroxyl (OH-) (S. Wang et al., 2020). Due to electrostatic attraction, these groups form complexes with NH4+, which promotes the uptake of this cation (Yin et al., 2019). Furthermore, the availability of metal elements in biochar enables the exchange of NH4+ with another cation which results in surface adsorption. Once NH4+ is adsorbed to the surface, it is no longer bioavailable for methanogens. Moreover, the porosity of biochar allows the colonisation and proliferation of microbial communities, particularly the methanogens within the pores. These pores offer shelter from NH3 stress during the AD process which increases the abundance of microbial communities in AD systems (G. Wang et al., 2022). Additional growth of microorganisms enhances the degradation of NH3-N in the AD system. For example, S. Wang et al. (2022) reported that the addition of rice straw biochar could increase the relative abundance of some bacteria and archaea such as Firmicutes, Bacteroidetes, Tenericutes, Synergistetes and Proteobacteria and Sedimentibacter, which enhanced the degradation of organic acids and ammonia–nitrogen in the anaerobic digestion system, resulting in an increase of biomethane production.
Biochar has been shown in previous reports to enhance the anaerobic digestion of CM (Ma et al., 2019; Ngo, Khudur, et al., 2022; Pan et al., 2019; Yu et al., 2021). However, different biochar feedstock types may not have a uniform effect on AD of chicken manure since the parent material from which biochar was derived has a dominant role in the determining the properties of the biochar (Joseph et al., 2021). While the effect of biochar on the AD of CM has been studied with biochar derived from wood, ricehusk, fruitwood, corn stover etc, no attempt has been made to investigate the effect of modified biosolids-derived biochar in the AD of CM (S. Wang et al., 2020; Yin et al., 2018; Yu et al., 2021).
Biosolids are by-products of the wastewater treatment process and their transformation to biochar and subsequent use in the AD of chicken manure would be of environmental benefit and may provide new management pathways for the wastewater industry. The amount of biosolids produced annually is high and is projected to increase in the future due to population growth and urbanisation (Roman-Perez et al., 2021; Wijesekara et al., 2016). In 2021, Australia produced 349,000 tonnes of dry biosolids which was 16% higher than the biosolids’ production in 2010 (300,000 tonnes) (Australian Water Association, 2021). The presence of contaminants such as per- and poly-fluoroalkyl (PFAs) substances and heavy metals in biosolids threatens their land-application in agriculture, which is the largest end-user of biosolids (Hakeem et al., 2022). Pyrolysis of biosolids would be a solution as it produces a useful biochar with reduced levels of PFAS and other contaminants (Thoma et al., 2022). In addition, biosolids-derived biochar contains micronutrients such as Co, Ni, Mo, and Fe, which play important roles in enhancing the microbial growth (Ippolito et al., 2020; Schattauer et al., 2011).
The properties of biochar could have an influence on its efficacy in anaerobic digestion and NH3 inhibition (Cai et al., 2022). Modification of biochar also provides an opportunity to improve the properties of biochar for enhanced AD. Several studies have demonstrated that acid and/or alkali modification of biochar increases the porosity and surface adsorption of biochar. For example, Vu et al. (2017) reported that the number of surface carboxyl and sodium-containing functional groups of corncob-derived biochar increased after the modification of the biochar with HNO3 (6 M) and NaOH (0.3 M). This caused an increase in the surface adsorption capacity of the biochar and the subsequent reduction of ammonia in the system. In another study, the modification of corn stalk and rice hull biochar with 0.2 mol L-1 H2SO4 increased the biochar adsorption capacity by 1.57 times, relative to the untreated biochar (Chen et al., 2021). This modification enhanced the electrostatic attraction between NH4+ and the surface functional groups of treated biochar, resulting in an increase in CH4 production. Modification of biochar aimed at improving the surface area and pore volume of biochar could be of relevance to increase the surface area and pore volume of biosolids-derived biochar.
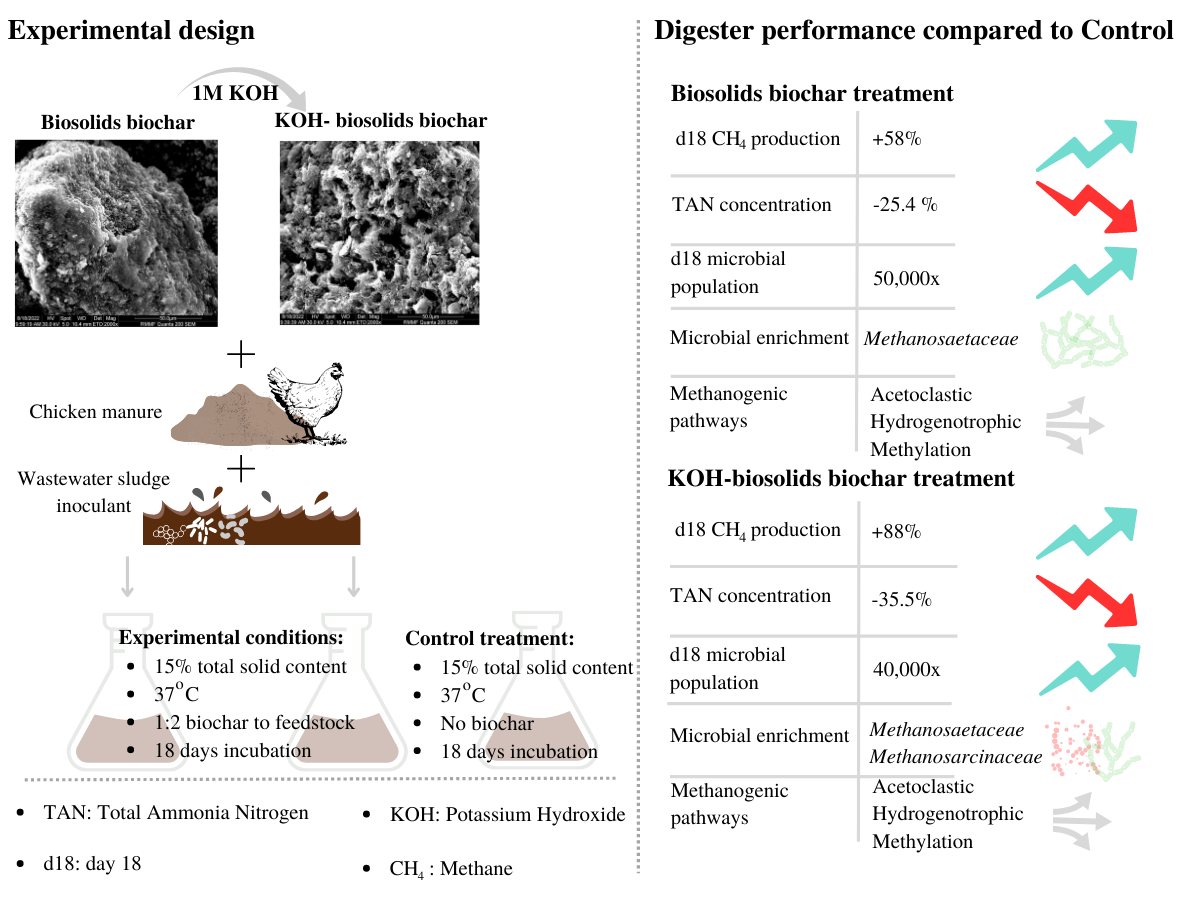
The overall aim of this present study is to examine the impact of pristine and KOH-modified biosolids-derived biochar on the anaerobic digestion of chicken manure. The specific objectives were to assess: (i) the effect of pristine and modified biosolids-derived biochar on biomethane production; (ii) the efficiency of biosolids biochar in reducing the level of Total Ammonia Nitrogen (TAN) in digesters; (iii) the effect of biochar on the microbial (bacterial and archaeal) community structure and population; (iv) the impacts of biochar modification on the anaerobic digestion process.
{kind=link}