Aedes albopictusis the predominant species in the Medellín Botanical Garden
From January 2018 – December 2019, adult and premature mosquitoes were collected every two weeks in the Medellín Botanical Garden. At four sites that had consistent water reservoirs (Fig. 1D), we collected 7376 larvae (5591 of which survived to adulthood). Aedes albopictus was the predominant species collected, accounting for 86.1% of the total larvae, followed by Culex spp. (13.9%) and Ae. aegypti (5.8%) (Table 1). Slightly more male larvae were collected, although both sexes were found in similar proportions (Table 1). Adults were captured at 19 different sites (Fig. 1D); 1398 adults were captured in total. Aedes albopictus was the predominant species (94.6%) followed by Ae. aegypti (4.2%). Culex spp. adults were rarely captured (0.14%). We were unable to identify 1.07% of the adults (Table 1). More adult males (of each Aedes species) were collected overall (Table 1).
Spatial distribution ofAedes aegyptiandAedes albopictusin the Medellín Botanical Garden
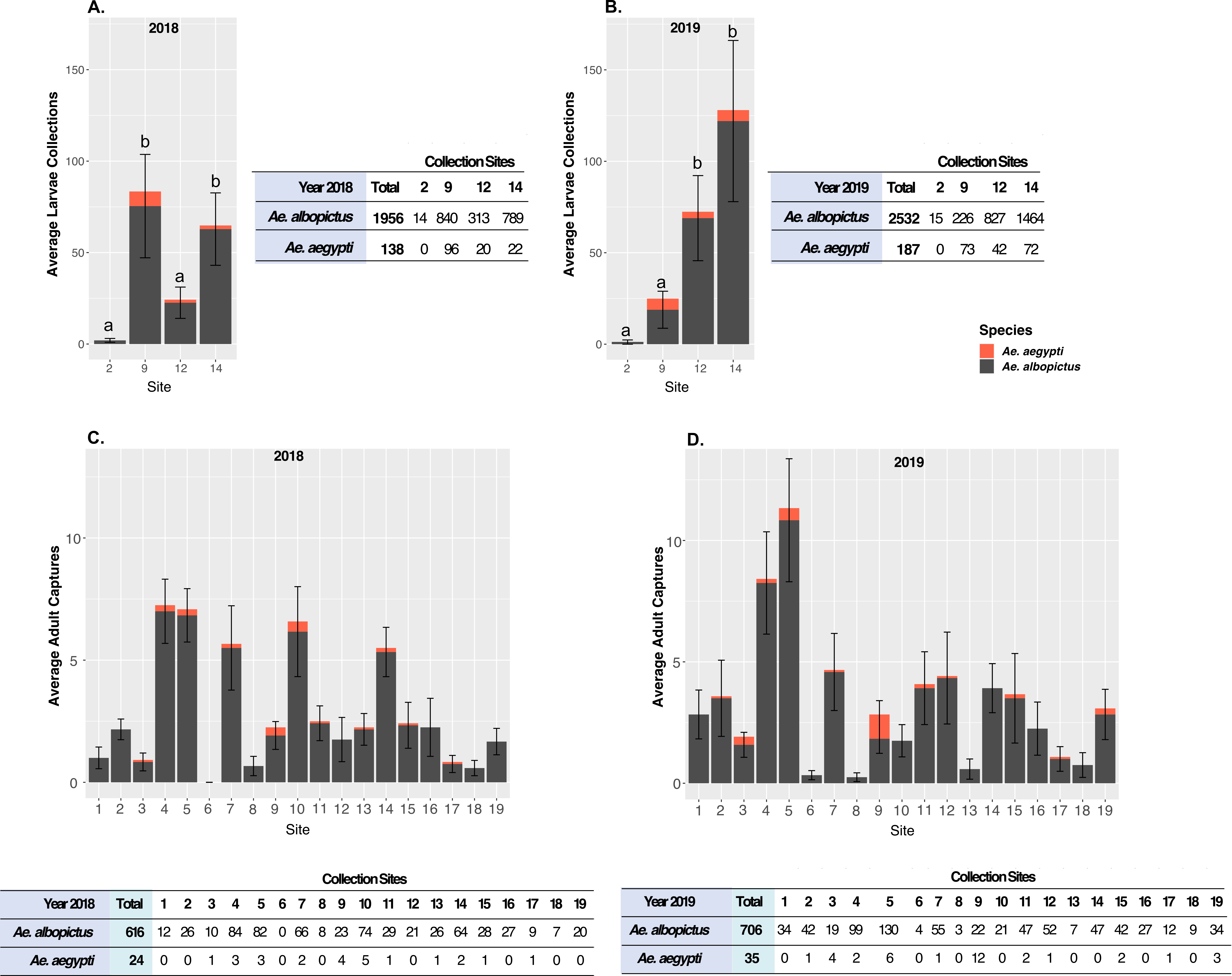
Because collection sites were variable (i.e., differences in vegetation, canopy cover, proximity to buildings; Table S1), we examined the spatial distribution of Ae. aegypti and Ae. albopictus within the park and determined exclusion or association sites of both populations. We found significant differences in the yearly average of larvae collected at each collection site between 2018 and 2019 (LMM: DF = 3, F = 5.7, p = 0.041; Figure S1A, B). The majority of larvae was collected at bamboo posts and tree holes: sites 9 and 14 (44% and 38% of the total larvae, respectively) in 2018, and at sites 12 and 14 (31.96% and 53.84%) in 2019. Regarding Aedes species, most Ae. aegypti larvae was collected at site 9 (69%) in 2018, and at sites 9 and 14 (39% and 38.5%, respectively) in 2019; most Ae. albopictus larvae was collected at sites 9 and 14 (42.9% and 40%) in 2018, and at sites 12 and 14 (57.8% and 37.6%) in 2019 (Figure S1A, B).
In our adult collections, no significant differences were observed between the yearly average of each site (LMM: DF = 18, F = 1.42, p = 0.11; Figure S1C, D), although we observed significant differences in population sizes between sites (LMM: F = 6.802, DF = 18, p < 0.001; Fig. 2A, B; Figure S1C, D). Aedes aegypti individuals were captured at eleven sites (Figure S1C, D), with most individuals captured at sites 4, 5, and 9 (Fig. 2A, Figure S1C, D; Table S3). Aedes albopictus were collected at all 19 sites; more than 60% were collected at sites 4, 5, 7, 10 and 14 (Fig. 2B, Figure S1C, D; Table S3).
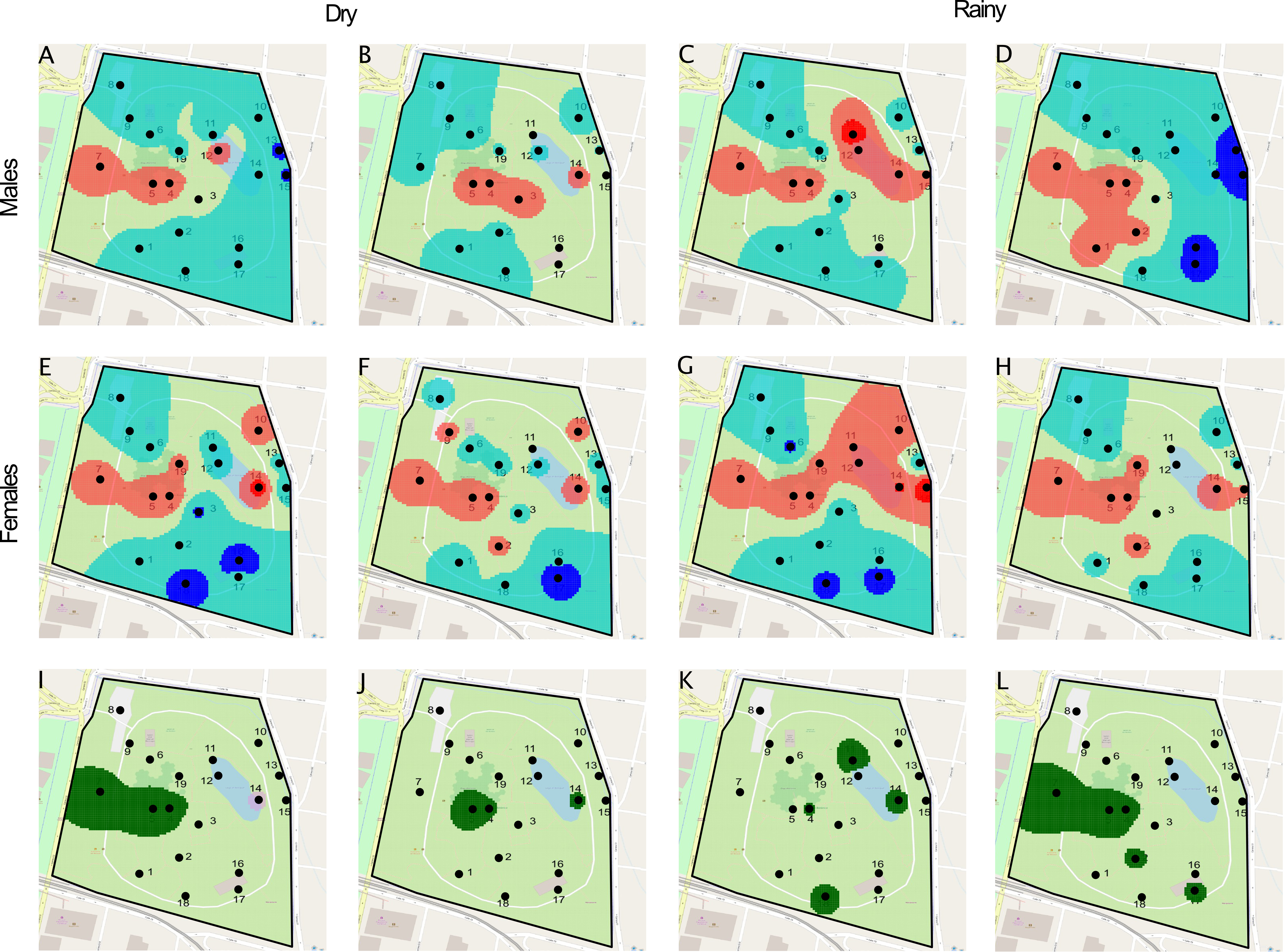
We next characterized the spatial aggregation of individuals of each species, and the spatial association between Ae. aegypti and Ae. albopictus using SADIE [30, 31]. The overall index of aggregation was Ia = 1.15 (Pa = 0.20) and Ia = 1.06 (Pa = 0.33) for Ae. aegypti and Ae. Albopictus, respectively, which suggests an overall moderate patchiness of both species across the sampled region that is not significantly different from a random pattern. However, we identified individual sites where populations aggregate, forming significant patches and gaps for each species (Fig. 2C, D). This departure of overall aggregation (Ia) from local indices (vi) may be a result of small sample sizes or edge effects (i.e., large or small counts consistently around the sampling area)—as in our case—when local indices are more powerful at detecting nonrandom distributions [35, 36].
We observed Ae. aegypti aggregations at five sites and a patch with significantly above-average density at site 4 (vi > 1.5; Fig. 2C). There were nine areas with low densities or gaps, while sites 14, 15 and 17 had a significantly below-average densities (Fig. 2C). A different local pattern was found for Ae. albopictus: we found aggregations at seven sites with a significantly above-average cluster at site 11 (vi > 1.5; Fig. 2D). There were eight sites with low density and a gap at site 17 (vj = -1.5; Fig. 2D). Spatial association between Ae. albopictus and Ae. aegypti was significant at sites 4 and 5 (p < 0.05; Fig. 2E), where both species aggregate and the majority of adults were captured (Fig. 2A, B; Figure S1C, D; Table S3). Significant local disassociations of Ae. aegypti and Ae. albopictus were found at sites 9, 11 and 14 (p < 0.05; Fig. 2E); only Ae. aegypti was aggregated at site 9 and only Ae. albopictus was aggregated at sites 11 and 14. Although we observed co-existence and exclusion locally, we did not observe significant disassociation (p = 0.5844) or association (p = 0.415) for the overall population across the study area.
Spatial distribution of male and femaleAedes albopictusin the Medellín Botanical Garden
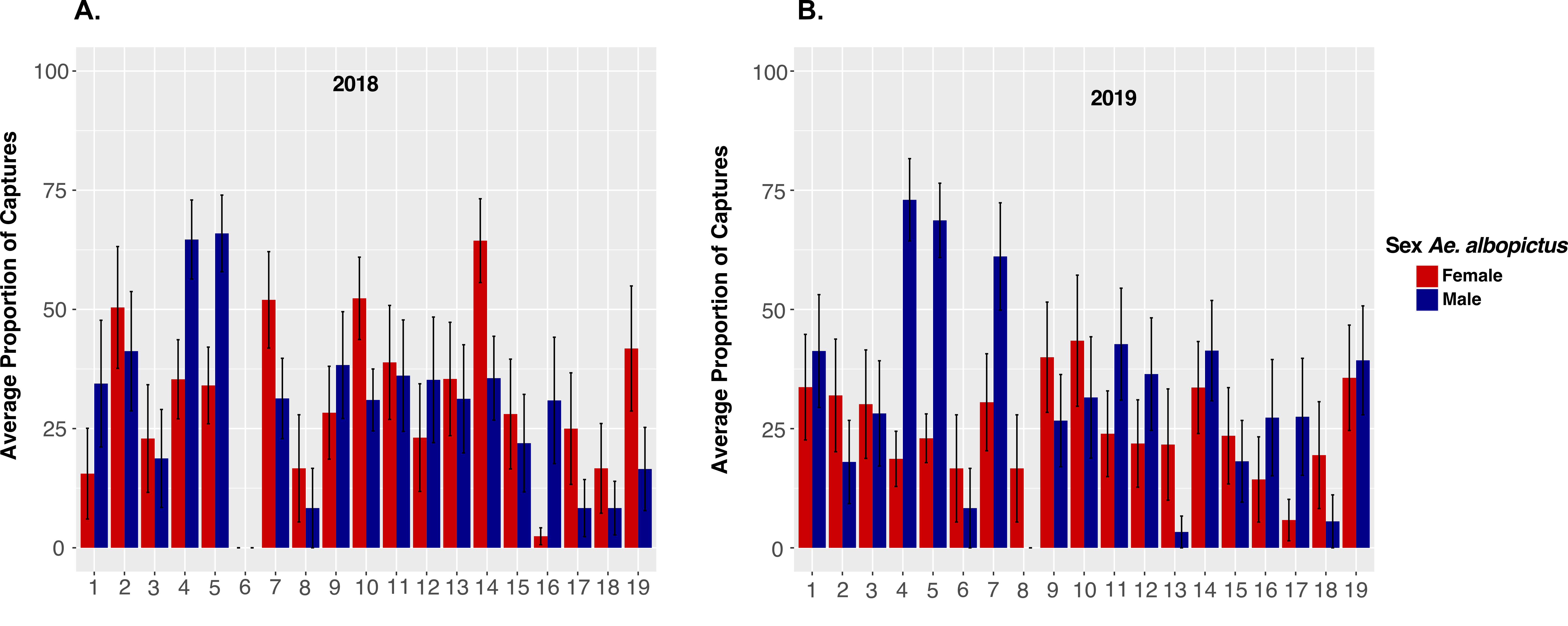
As we collected more Ae. albopictus males than females during this study (58.4% vs. 41.6%; Table 1), we examined Ae. albopictus male and female distribution within the park to identify areas where they aggregate or disperse. The average proportion of males and females collected at each site did not significantly differ between 2018 and 2019 (GLMM: DF = 18, F = 0.654, p = 0.1; Figure S2). However, we observed significant differences of the average male-female proportions between sites (GLMM: F = 2.7041, DF = 18, p < 0.001; Fig. 3A, B). The largest proportion of males were collected at sites 4 and 5 (Fig. 3A; Figure S2), two sites with a high vegetation density (Fig. 1, Table S1). Females had higher proportions at sites 10 and 14 (Fig. 3A, B), sites with low and high vegetation, respectively.
The overall distribution of males and females was moderately patchy, but not significantly different from what is expected by chance (Males: Ia = 0.9, Pa = 0.63; Females Ia = 1.18, Pa = 0.157). Locally, however, we found that males aggregated in fewer patches than females (Fig. 3C). Additionally, there is a cluster with a high density of females at site 15 (vi > 1.5; Fig. 3D). Both sexes had low densities at seven sites. However, sites 2, 10 and 19 were occupied by females but not by males. Significant gaps for females were found at sites 16 and 17 (vi < -1.5; Fig. 3D). We found that both sexes were significantly associated across the sampled area (p = 0.016). Significant local associations were observed at sites 5, 7 and 11 (Fig. 3E), where high numbers of both males and females were recorded (Fig. 3A, B). Site 17 also had a significant local association, due to concomitant small counts of both sexes (Fig. 3C, D).
We further analyzed the spatial distribution of males and females during the dry and rainy seasons of Medellín. Medellín has two distinct periods of high and low precipitation annually (see Fig. 5). We combined data for the dry seasons (first: January and February; second: June, July and August) and rainy seasons (first: March, April and May; second: September, October, and November). At a local scale, we found a similar pattern of patch and gap distribution for both males and females during the first and second dry season (Figure S3A, B, E, F). However, the distribution of patches and gaps for both sexes differed between the first and second wet season. Interestingly, the pattern observed in the second wet season resembled that observed in the first dry season for both sexes (Figure S3C, D, G, H). Due to the within-year variation observed for the rainy and dry seasons, it is difficult to describe general differences in Ae. albopictus distribution for both sexes. Differences in male and female distribution in the dry and wet seasons may indicate that each sex aggregates differently in space during the year and that this distribution may be influenced by weather variables. We also observed significant associations in certain areas of the park of both sexes, mainly at sites 4 and 5 (Figure S3I, J, K, L) where high vegetation was found.
Vegetation coverage influencesAedes albopictuscaptures
We next examined if vegetation coverage influenced Ae. albopictus captures, classifying each collection site by its percentage vegetation cover, which ranged from very low to very high (Table S1). Vegetation coverage had a significant effect on adult captures (LMM: F = 2.8583; DF = 4; p < 0.001; Fig. 4A). The majority of Ae. albopictus were captured at areas classified as high or very high vegetation coverage (Fig. 4A). Males and females were uniquely distributed across the differing vegetation coverages (LMM: F = 16.7731; DF = 4; p < 0.001; Fig. 4B)—significantly more males were found at areas with very high or high vegetation coverage sites compared to females, who were similarly distributed between sites with low to high vegetation coverage (Fig. 4B).
Aedes albopictushave a bimodal distribution in relation to weather variables
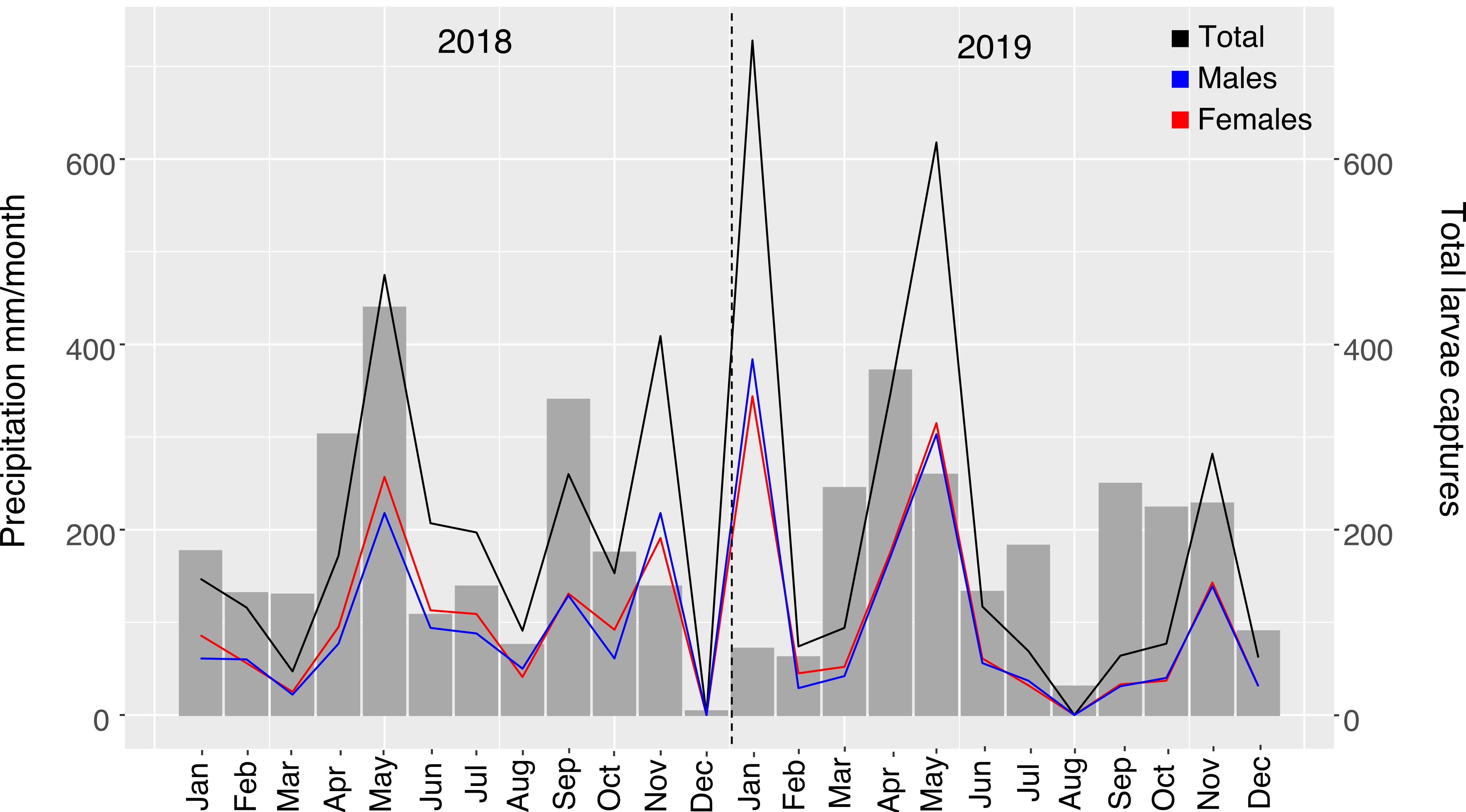
We examined how weather variables correlated with our monthly Ae. albopictus collections. Aedes albopictus captures had a bimodal distribution with distinct peaks during April-May and October-November in both 2018 and 2019, coinciding with months with the highest cumulative precipitation (Fig. 5A); we found a significant positive correlation with male and female captures during these months (LMM: p < 0.05; Table S4). Relative humidity also had a significant positive correlation with male and female captures (LMM: p < 0.05; Fig. 5B; Table S4). Temperature and wind speed showed a significant inverse correlation with adult captures (LMM: p < 0.05; Fig. 6B; Table S4). We also observed a significant correlation between precipitation and total larvae collected (LMM: F = 10.19; p = 0.0042; Table S5)—the highest number of larvae collected occurred during months with the highest precipitation in both 2018 and 2019 (Figure S4), although we also collected larvae in high numbers in January of 2019, a month with low rain levels. We found no significant correlation with the total larvae collected for other evaluated environmental variables (Table S5).
Environmental factors influenceAedes albopictusbody size
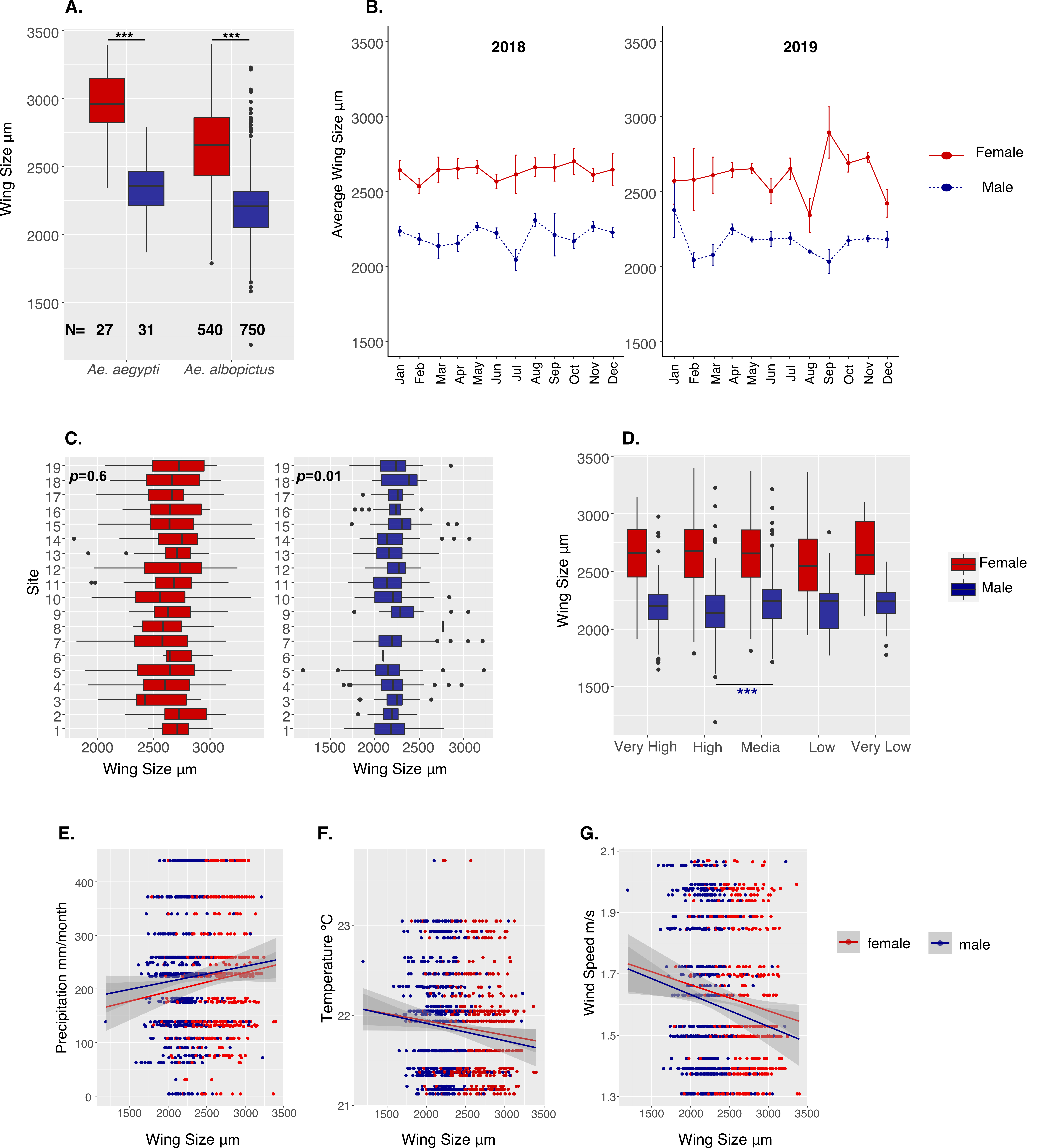
Because mosquito body size is related to female longevity and reproductive output [37, 38], we examined how Ae. albopictus size changed during the study period and asked if collection site or environmental variables influenced this trait. Using wing length as a proxy for body size [29], we found significant differences in size between species and sex of the adults collected (LMM: DF = 1, F = 4.71, p = 0.030; Figure S5A). Female Ae. albopictus had an average wing length of 2638.16 ± 0.55 µm and males 2197.18 ± 0.31 µm. Female Ae. aegypti had an average wing length of 2914.54 ± 11.26 µm and males 2335.65 ± 7.73 µm (Figure S5A).
We next analyzed how size changed during the study period by testing the significance of month, collection site, environmental variables and size as predictor variables of Ae. albopictus captures. We found that male size significantly changed with the month during the study (LMM: DF = 5, F = 2.64, p = 0.022; Figure S5B) but female size did not (LMM: DF = 5; F = 1.309; p = 0.258). Similarly, we found an effect of collection site (LMM: DF = 18; F = 1.855; p = 0.01; Figure S5C) and vegetation coverage on male size (LMM: DF = 4; F = 3.97; p = 0.003; Figure S5D)—larger males were found in sites with medium vegetation coverage. In contrast, female size was not affected by collection site (LMM: DF = 18; F = 1.894; p = 0.265; Figure S5C) or vegetation coverage (LMM: DF = 4 ; F = 1.491; p = 0.20; Figure S5D). We also observed a significant association of Ae. albopictus size with precipitation, temperature and wind speed for both sexes (Table S6). Precipitation was directly proportional (Figure S5E), while temperature and wind speed were both inversely proportional to adult wing size (Figure S5F, G).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}