RNA sequencing and taxonomic annotation
A total of 263 059 818 raw reads were obtained from 24 RNA samples which eventually passed quality control, with an average of 10 960 826 reads per sample. After merging and the removal of chloroplasts and mitochondria reads, we obtained a total of 15 029 taxonomic units from our RNA-Seq library from which 1 059 of the taxonomic units were filtered out due to low relative read numbers abundance (read number mean abundance < 10⁻5). The microbial diversity and community structure were thus analyzed based on a total of 9 594 prokaryotic and 4 376 taxonomic units were assigned to the Eukaryota (Supplementary table 1).
With our original experimental design, we aimed for a total of 38 pooled RNA samples obtained as duplicates from five time points in time (T1 = 2 days, T2 = 7 days, T3 = 14 days, T4 = 21 days and T5 = 53 days). However, due to low RNA yield from the biofilm specimen, only 24 samples (none of T4 = 21 days) passed the RNA sequencing quality control with an average amount of 28.32 ng RNA per sample from four sampling points in time.
Soil and time dependent biofilm succession on PE and glass
We tested whether we could observe different stages of biofilm succession on soil-buried PE foil and performed a PERMANOVA analysis (Table 1) in order to identify the different conditions shaping this biofilm community. To do so, we used the taxonomic composition assigned from our RNA transcripts and calculated their relative abundance within the active portion of the sequenced community to calculate a PCoA plot based on Bray-Curtis measures of dissimilarity (Fig. 1).
Table 1
PERMANOVA testing of factors explaining differences in biofilm community
| | df | Sums of square | Pseudo F | R² | P value | Significance |
| PE vs. glass | 1 | 0.4566 | 1.2581 | 0.05 | 0.266 | - |
| Forest soil vs. Plastic soil | 1 | 1.8062 | 5.9899 | 0.21 | 0.001 | *** |
| Days | 3 | 2.0328 | 2.1151 | 0.24 | 0.002 | ** |
| Early biofilm (Day 2 vs. rest) | 1 | 0.5118 | 1.4202 | 0.06 | 0.152 | - |
| Early mid biofilm (Day 7 vs. rest) | 1 | 0.7724 | 2.2162 | 0.09 | 0.015 | * |
| Late mid biofilm (Day 14 vs. rest) | 1 | 0.3082 | 0.8338 | 0.04 | 0.584 | - |
| Late biofilm (Day 53 vs. rest) | 1 | 1.1902 | 3.6116 | 0.14 | 0.001 | *** |
According to PERMANOVA analysis, the type of soil explained 21% of the observed differences between the biofilm communities (Table 1), whereas the type of substrate (PE vs. glass) did not contribute to a significant shift of the biofilm community structure (R² = 0.05, p = 0.266).
We observed a clear separation of the sequenced biofilm communities incubated in plastic soil versus forest soil, which deviated along the primary principal axis (Fig. 1). According to PERMANOVA analysis, the type of soil explained 21% of the observed differences between the biofilm communities. Within the two soil types, the most apparent separation of the microbial communities was explained by the different successional stages of the biofilm according to days of incubation (Fig. 1). In both plastic soil and forest soil, the microbial communities changed along the progression of the biofilm from an early biofilm (Day 2) to a mid- (Day 7) and late biofilm succession (Days 14–53). The most advanced point of our biofilm incubation experiment (Day 53) showed also the most distinct microbial beta diversity, which differed significantly from the rest of the sequenced time points in both forest- and plastic soil (PERMANOVA: R² = 0.146, p = 0.0012).
Most striking was the community succession of the incubated biofilm in plastic soil: All four time points formed individual clusters along the two axes of the PCoA (Fig. 1) and a development of the biofilm communities was observed based on their taxonomic shifts from early- to mature colonization. In contrast to that, the community structure of the biofilms in forest soil clustered closely together during the early- and mid-biofilm successional stages and showed a significant shift in community structure only on day 53 (Fig. 1).
To test the activity and function of the biofilm microbial taxa, we compared their expression levels of 16S/18S rRNA gene transcripts for taxonomic assignment (Fig. 1–3) at the given points in time as well as the expression levels of specific genes relevant f or the PE-plastisphere (Supplementary table 2) for functional profiling (Fig. 4–5). By using this metatranscriptomic approach, we were able to follow only the active portion of the PE-associated plastisphere community during their biofilm succession over 53 days in the two different soils.
Plastic soil
The PE-associated biofilm communities in plastic soil developed over the course of time into a less diverse community compared to their initial successional stage (Fig. 2). After an initial increase in diversity at day 7 (MedianChao1 = 4732.7, MedianH’ = 5.42, MedianD’ = 0.99), the microbial diversity decreased significantly within the biofilm of the late successional stage at day 53 (MedianChao1 = 3899.8, MedianH’ = 2.85, MedianD’ = 0.78 ). We found no significant differences in alpha diversity between the biofilm community on PE vs. glass at any successional stage. Concerning the taxonomic makeup of the attached microbial communities, however, we did observe slight differences between the microbial community attached to PE when compared to the biofilm community on glass (Fig. 2). A preference of known plastic-associated taxa (e.g Rhodococcus, Nocardioides and Streptomyces) to PE surfaces was observed in the mid-biofilm stages as compared to glass, however, none of them were exclusively active on PE (Fig. 2).
In the early- to mid-biofilm stages (between 2 and 7 days), transcripts of the Actinobacteriota and Firmicutes were the most abundant prokaryotic phyla of the biofilm communities in plastic soil (Fig. 2). Transcripts of the genera Lamia (4.9% of mean relative abundance), Mycobacterium (3.3%), Nocardioides (2.2%), Rhodococcus (2.4%) and Streptomyces (2.0%), were dominant in the early stages of biofilm succession, followed by the members of the Firmicutes phylum Sporosarcina (1.4%) and Bacillus (1.1%). All of them decreased in their SSU rRNA abundance during biofilm succession (Fig. 2) and disappeared in the late biofilm stage (e.g Proteobacteria) or were otherwise present in abundances below 0.1% (Firmicutes). Other present SSU rRNA transcripts in early biofilm succession were Chloroflexia JG30-KF-CM45 (2.0%) of the Chloroflexi and Pedosphaeraceae of the Verrucomicrobiota phylum (1.2%). Proteobacteria were represented by transcripts of the genera Amaricoccus (0.7%), Mesorhizobium (0.8%) and Rhodobacteraceae (0.6%).
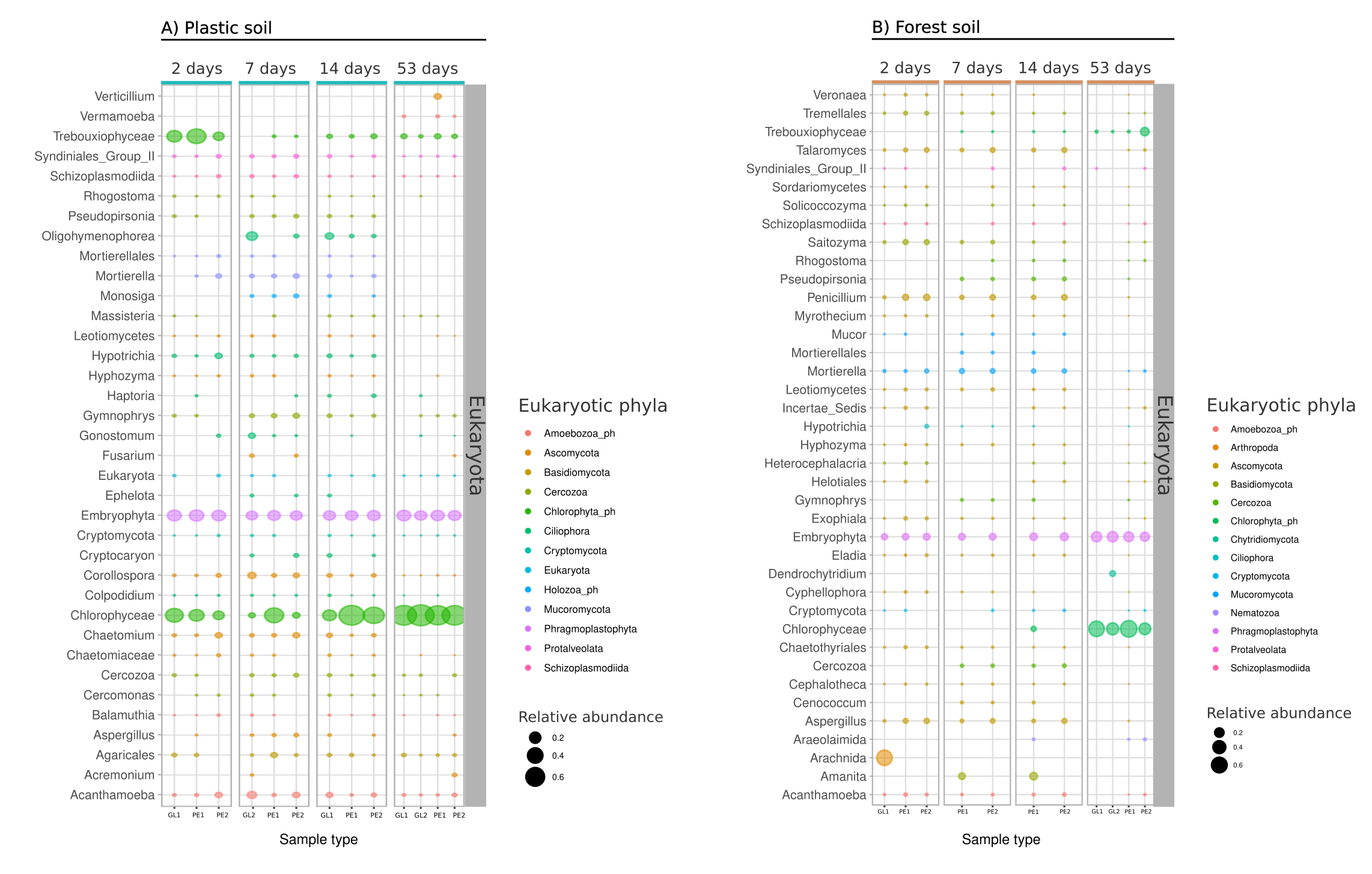
The active eukaryotic fraction of the biofilm community consisted of the Ascomycota Aspergillus (0.9%), single-celled protists like Cercozoa (1.0%) and Cercomonas (0.3%) (Supplementary Figure S1 A). Other abundant Eukaryotic members of the community were the saprotrophic fungi Mortierella (1.8%), as well as the protozoa (Ciliophora) and the order of Agaricales (gilled mushroom). Trancriptomic reads originating from Embryophyta (plants) and Acanthamoeba (amoeba) were likewise present across all communities (Supplementary Figure S1 A).
Moving further in time, the most apparent shift in activity of the microbial community was detected between Day 14 and 53 of biofilm succession. The transition from a mid-succession stage to a late biofilm community was accompanied by a strong increase of Cyanobacterial transcripts (up to 49% of all sequenced reads per sample) and a decrease of other early-to mid-biofilm members (Fig. 2). Highly abundant were autotrophic genera of the Nostocaceae, e.g Nostoc_PCC and Tolypthrix. Other eukaryotic late colonizers were the green algae Chlorophycae (7%) and Trebouxiophycae (2.5%) of the Chlorophyta phylum (Supplementary Figure S1). Firmicutes members of the late biofilm succession were Bacillus (1.0%) and Sporosarcina (0.9%), whereas the Chloroflexi phylum was represented by the genus A4b of the Anaerolineae (1.3%), JG30-KF-CM45 (0.6%) and Caldilineaceae (0.5%). Also, phylum members of the Planctomycetota were a dominant part of the late biofilm community with Gemmataceae (1.24%), SM1A02 (1.2%) and Pirellulaceae (0.67%). From the Actinobacteriota only Microbacterium (1%) and Microtrichaceae (1.5%) were present at that late succession stage. Higher abundant eukaryotic transcripts were again derived from Embryophyta (17%) and reads of the grazing protists Acanthamoeba (1%) and Vermamoeba (0.6%).
Forest soil
Similar to the biofilm communities in plastic soil, we saw a first slight increase in diversity peaking at day 7 of incubation (MedianChao1 = 4279 MedianH’ = 4.14, MedianD’ = 0.94) and a gradual drop in diversity towards the late successional stage after 53 days (Fig. 3). The differences in forest soil where, however, smaller than in plastic soil and none of them statistically significant (Fig. 3). The sequenced biofilm community in forest soil stayed overall more stable during biofilm succession, the taxonomic differences were also less distinct between the different biofilm stages (Fig. 3).
The early biofilm community was again dominated by Actinobacteriota such as Acidothermus (32.9%), Mycobacterium (6.9%), Conexibacter (3.5%) and Acidimicrobiia (3.6%), which where the top initial colonizers in the biofilm community on both substrates (Fig. 3). However, the early biofilms derived from forest soil contained more members of the Proteobacteria, such as Elsterales (5.9%), Acetobacteraceae (1.9%) and Roseiarcus (1.6%), as well as members of the phylum Planctomycetota, e.g the cellulose degrader Gemmataceae (2.6%).
Compared to plastic soil, there were less taxonomic changes during biofilm succession until day 53 of incubation in forest soil. During the late succession stage of the biofilm, there was a tremendous decrease in taxonomic richness on glass compared to that of PE (Fig. 3 and Figure Supplementary Figure S1 B). Both the Bacterial- and Eukaryotic fraction of the biofilm on glass consisted of a few highly abundant taxa: The Actinobacteriota Acidothermus was highly abundant with up to 43.4% of all reads in a given sample. Equally abundant were the genera Gemmataceae and Pirellulaceae of the Planctomycetota phylum. A reduced taxonomic diversity on glass-associated biofilm was mostly visible in the top 50 prokaryotic taxa (Fig. 3), as only seven of the top genera were present in the mature biofilm (day 53) on glass compared to the 31 unique genera of higher abundance present within the PE-associated biofilm of day 53.
Concerning the eukaryotic fraction of the biofilm community, fungi were more abundant in forest soil than in plastic soil. The genera Aspergillus (3.4%) and Penicillium (5.3%) of the phylum Ascomycota were abundant in the early- to mid-successional stages of the biofilm community, yet decreased in their abundance within the mature biofilm community (Supplementary Figure S1 B). Contrasting to the eukaryotic community in plastic soil, we detected no members of the Chlorophyta phylum (green algae) in the early- to mid-biofilm stage but a strong domination of the late biofilm stages by these genera (Supplementary Figure S1 B). The changes in bacterial composition and activity were less pronounced compared to plastic soil. In addition, no Cyanobacteria were observed in the forest soil biofilm community.
PE-specific gene expression marks successional biofilm stages on PE and glass
In search of specific pathways during the succession of PE-associated biofilms, we used our KEGG annotated metatranscriptomic data and compared it to a set of 35 known enzymes involved in PE- and/or general polymer colonization (Figs. 4 and 5), five nif genes related to nitrogen fixation (Fig. 6) as well as 133 biofilm related enzymes involved in lipopolysaccharide biosynthesis and extracellular matrix synthesis, respectively (Supplementary Figure S2). Gene expression patterns of the analysed PE-specific genes varied over the course of biofilm succession. Some of the involved pathways showed an overexpression only at specific successional stages, which we verified in detail by DESEQ2 analysis of differential abundance (Supplementary table 3). In general, we detected a later onset of PE-specific pathways in forest soil when compared to the same genes in plastic soil biofilm communities (Fig. 4A and 4B).
During the early colonization in plastic soil (day 2–7) of PE foils, we found a high activity of genes involved in fatty acid degradation while in forest soil this occurred later during the mid-successional stage (day 7–14). For example, transcripts of the acyl-CoA dehydrogenase (ACADM), a putative acyltransferase (atoB), 3-ketoacyl-CoA thiolase (FadA) and a fatty acid oxidation complex subunit alpha (FadJ) were expressed and overly abundant in this initial biofilm stage on PE foil in plastic soil (Fig. 4A). All of these genes are involved in either aerobic long-chain fatty acid degradation (fadB and fadA), or the anaerobic equivalent of the same pathway (fadJ). Even though transcripts of atoB and FadA were detected in one glass sample at day 2, fatty acid degradation was less prevalent on glass than on PE foil in plastic soil for the rest of the biofilm stages (Fig. 4A).
The initial biofilm stage on day 2 was also shaped by genes encoding membrane transporters such as the ATP-binding proteins (ABC transporters) LivF, LivH, LivM and LivG which were upregulated in both glass- and PE biofilm communities in forest- and plastic soil (Fig. 4A and 4B). The long-chain fatty acid transporter FadL and the TRAP-like transport protein yiaO, both involved in the transport of aliphatic hydrocarbons, were active in two initial biofilm samples on PE foil in plastic soil during day 2 and day 7, as well as in a mature PE-associated biofilm sample of day 53 (Fig. 4A). In forest soil, these two transporters were active only in the biofilm samples of day 7–14 (Fig. 4B).
Genes involved in the degradation of n-alkanes were the alkane-monooxygenase (alkM), the S-(hydroxymethyl)glutathione dehydrogenase (frmA), an aldehyde dehydrogenase (ALDH) and the long-chain alkane monooxygenase (ladA), all of which were also active during the early successional stages (day 2–7) of PE-associated biofilm from plastic soil (Fig. 4A) and a bit later (day 7–14) in biofilm communities from forest soil (Fig. 4B). The expression of genes related to n-alkane degradation were therefore highest in early- to mid-successional stages and dropped in activity towards the late biofilm communities. Other enriched pathways were genes involved in the breakdown of secondary plant metabolites and natural terpenes, such as geraniol and citronellol (atuC,atuD and atuF), all of which were active during day 7–14 in forest soil and during the initial successional stage (day 2) in plastic soil (Fig. 4). The only exception was the related isohexenylglutaconyl-CoA hydratase (atuE), which was active exclusively during the late successional stage of day 53 in both soils (Fig. 4).
The mid successional stage in forest soil was characterized by the expression of enzymes involved in fatty acid beta-degradation, such as fadJ (3-hydroxyacyl-CoA dehydrogenase), fadB (Fatty acid oxidation complex subunit alpha) and ACADM (acyl-CoA dehydrogenase), which had their peak of expression on day 14 (Fig. 4). Also, genes from the pathways of geraniol degradation were overexpressed during this phase of biofilm development. Noteworthy in the context of possible PE-degrading pathways, was the overexpression of the extracellular lipase lip, as well as the Acyl-CoA dehydrogenase DCAA, capable of degrading caprolactam – the monomer of Nylon 6. Moreover, we found almost all genes involved in the N-alkane degradation pathway to be overly active at certain biofilm stages (Fig. 4). Identical to genes expressed in plastic soil, the Alkane 1 monooxygenase alkM (candidate for PE-degradation) was active during day 7 and day 14 of biofilm formation and remained downregulated during both the initial biofilm phase (day 2) and the late biofilm succession (day 53). To our surprise, we found the polyethylene terephthalate degrading enzyme PETase to be present and expressed by the biofilm community in their mid-successional stage in forest soil, yet entirely absent in plastic soil.
Overall, the late biofilm samples (day 53) showed only a few genes to be expressed above the average (Fig. 4). With the exception of a single PE sample in plastic soil (Fig. 4A, PE1 day 53) we observed an overall decline in the expression of genes relevant for PE-degradation towards the late successional phases of biofilm growth (Fig. 5). In fact, most genes that were highly active in the preceding biofilm stages were underexpressed in the mature biofilm communities. Out of genes involved in fatty acid beta-oxidation, only fadB and fadN (both Fatty acid oxidation complex subunit alpha) were slightly above the average in the late biofilm stage of plastic soil and forest soil, respectively (Figs. 4 and 5). Other earlier abundant enzymes, such as ABC transporter genes or the PE-specific candidate genes Alkane 1 monooxygenase alkM and the extracellular lipase lip were remarkably underrepresented in the late biofilm stage. Moreover, nonspecific genes involved in the biosynthesis of secondary metabolites (3-oxoacyl-[acyl-carrier protein] reductase FabG and a putative enoyl-CoA hydratase atuE) were overrepresented on day 53 of incubation in both soils. One sample of the biofilm community on PE foil (PE1day53), showed a remarkably high expression of PE-specific genes compared to other samples from the same sampling day (Fig. 4A). These highly expressed genes included ABC transporters (e.g LivH, LivF and LivG/CysA), as well as genes involved in fatty acid metabolism including the extracellular lipase lip (Lactonizing lipase precursor) and fadA (3-ketoacyl-CoA thiolase). In the forest soil, we observed a clear functional pattern that distinguished the different successional biofilm stages from each other in regard to PE-specific genes. (Fig. 4B). Especially genes encoding dehydrogenases like fadE (acyl-CoA dehydrogenase) and frmA (S-(hydroxymethyl)glutathione dehydrogenase) were considerably active compared to later biofilm stages. Unique for the sequenced PE biofilm communities in this early stage, was an overexpression of the extracellularly secreted carboxylic-ester hydrolase phaZ (Esterase PHB depolymerase) in PE communities compared to the biofilm on glass (Fig. 4).
Plastic soil microbial community showed high expression of nif genes to overcome nitrogen limitation on PE in the mature biofilm
Apart from genes related to PE degradation, we detected the expression of genes responsible for microbial nitrogen fixation (Fig. 6) in the biofilm community. Most striking was the increase in expression of a number of nif genes over the course of biofilm succession in plastic soil, which encode a variety of nitrogenases and are an important part in the microbial nitrogen fixation pathway (Fig. 6A).
In plastic soil, the expression of nifD, nifH, nifK, nifN and nifT was the highest in the mature biofilm stage, with nifH reaching the highest gene expression on PE foil after 53 days of incubation (Fig. 5A1). Transcripts of nifD, nifH and nifK were, however, present during the mid-successional biofilm stage on two PE samples (Fig. 6) and on a single glass sample on day 2 of incubation. The highest expression levels were reached on the PE samples at this stage of biofilm succession, even though the biofilm of a single glass sample also expressed four of the nif genes in low abundances. To our surprise, we could not detect a single transcript of nif genes related to nitrogen fixation in our forest soil biofilm community.
Biofilm succession and surface coverage using SEM image analysis
In addition to the transcriptomic data of the microbial biofilm communities on soil-buried PE foil, we used SEM image analysis to visually follow growth and successional development of the biofilm stages in both soils (Fig. 7 and Fig. 8). In order to quantify the surface coverage, we used a threshold-based image analysis which depicted biofilm structures from the background surface and calculated the extent of microbial coverage (Supplementary Figure S3).
An initial biofilm growth was visible on day 2 of incubation in both soil types. Over the course of time, we observed a higher surface coverage of biofilms in plastic soil (Fig. 7) compared to the biofilm growth in forest soil (Fig. 8). Biofilm complexity on the PE surface increased with time of incubation in both soils with a variety of morphological biofilm features. Structures of extracellular polymeric substances (EPS) were detected within the biofilm, where strings connected the thick microbial mats with the surface (Fig. 7: day 53).
We found various shapes of bacterial cells (e.g. singular cocci and rod-shaped bacteria as well as streptococci and tetrads) in close arrangements on the surface and embedded within surface grooves of the PE foil (Figs. 7 and 8). Most apparent were the large fungal hyphae structures from day 14 on and a number of fungal spores on the PE foil. In the late biofilm stage within the plastic soil, we found an increased number of filamentous bacteria as well as streptococci and chains of bacilli (Fig. 7: day 53). In general, the surface coverage of PE samples with biofilm in plastic soil was higher than those incubated in forest soil. The PE surface in plastic soil was covered in thick biofilm after 53 days of incubation, where mean coverage reached 39% of the PE surface (Fig. 7). Especially in the middle stages of biofilm growth (14 days), there were numerous bacterial cell aggregates visible on PE, as well as fungal structures like hyphae and spores which formed a substantial microbial layer on top of the surface. In forest soil, mean surface coverage was highest at day 14 with 2.3% (Fig. 8: day 14) and did not reach higher coverages throughout the incubation. Considering these low coverage numbers, we detected an earlier onset of biofilm development in plastic soil compared to the succession in forest soil similar to the patterns visible in the metatranscriptome data.
{kind=link}
{kind=link}