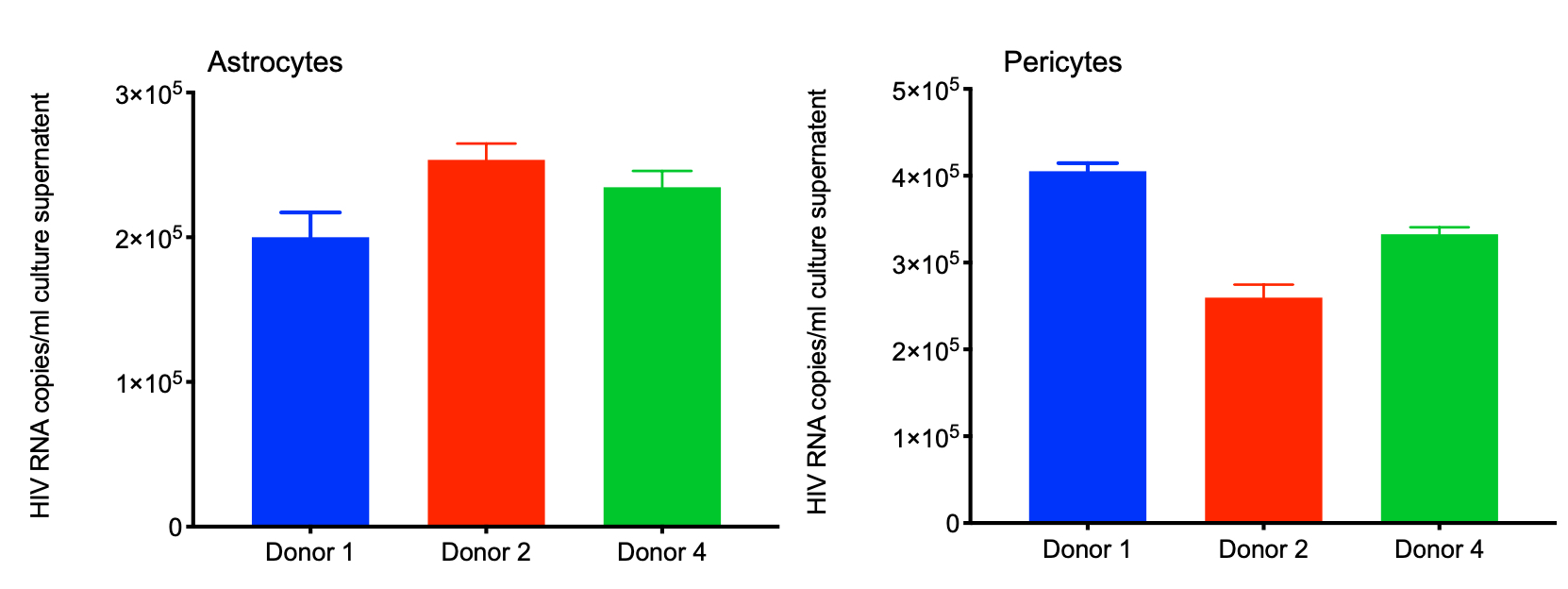
Neurocognitive impairments associated with COVID-19 are increasing, both during acute infection and in post-acute sequelae of COVID-19 (PACS). The impact of SARS-CoV-2 infection in the central nervous system (CNS) of people living with HIV (PWH), who already have compromised blood-brain barrier (BBB) and persistent brain inflammation, is not well studied. To address this, we investigated the consequences of HIV/SARS-CoV-2 coinfection in primary human astrocytes and pericytes, which are major targets of SARS-CoV-2. We found that SARS-CoV-2 replication was limited in healthy and HIV-infected astrocytes and pericytes. SARS-CoV-2 Mono and co-infection with HIV resulted in a modest increase in the expression of viral host cell entry factors and pro-inflammatory mediators like IL-6 and IL-1β in astrocytes. Using quantitative proteomic analysis, we identified a distinct protein expression signature in co-infected astrocytes/pericytes compared to healthy, SARS-CoV-2, or HIV-infected cells. The regulated pathways in co-infected cells are associated with the development of neurological disorders such as PD, AD, HD, and ALS. Several symptoms of these diseases are frequently observed in neuro-PASC patients [35]. In addition, these findings have critical implications for PWH, as they frequently experience symptoms resembling these neurological disorders [36].
Viral infections manipulate host cell machinery for their survival and immune evasion [37]. The expression of host cell entry factors plays an essential role in establishing and persistence of viral infection. We examined the expression of SARS-CoV-2 entry receptors in astrocytes and pericytes infected with the virus alone or in combination with HIV. Our findings align with previous studies including ours, showing low to moderate expression of ACE2 and NRP1 in healthy and SARS-CoV-2 infected brain cells [38–41]. Research using brain tissue from COVID-19 patients suggests that ACE2-expressing pericytes may facilitate SARS-CoV-2 entry into the brain [18]. Similarly, in cortical assembled models, pericyte-like cells expressing ACE2 serve as sites for viral replication [42]. Furthermore, studies on human monocyte derived macrophages have shown that restricted SARS-CoV-2 infection triggers dysregulated immune responses and overexpression of pro-inflammatory cytokines [43, 44]. Taken together, these findings support our observations that restrictive SARS-CoV-2 infection in healthy and HIV-infected astrocytes and pericytes leads to a modest increase in inflammatory mediator expression, potentially disrupting normal brain function.
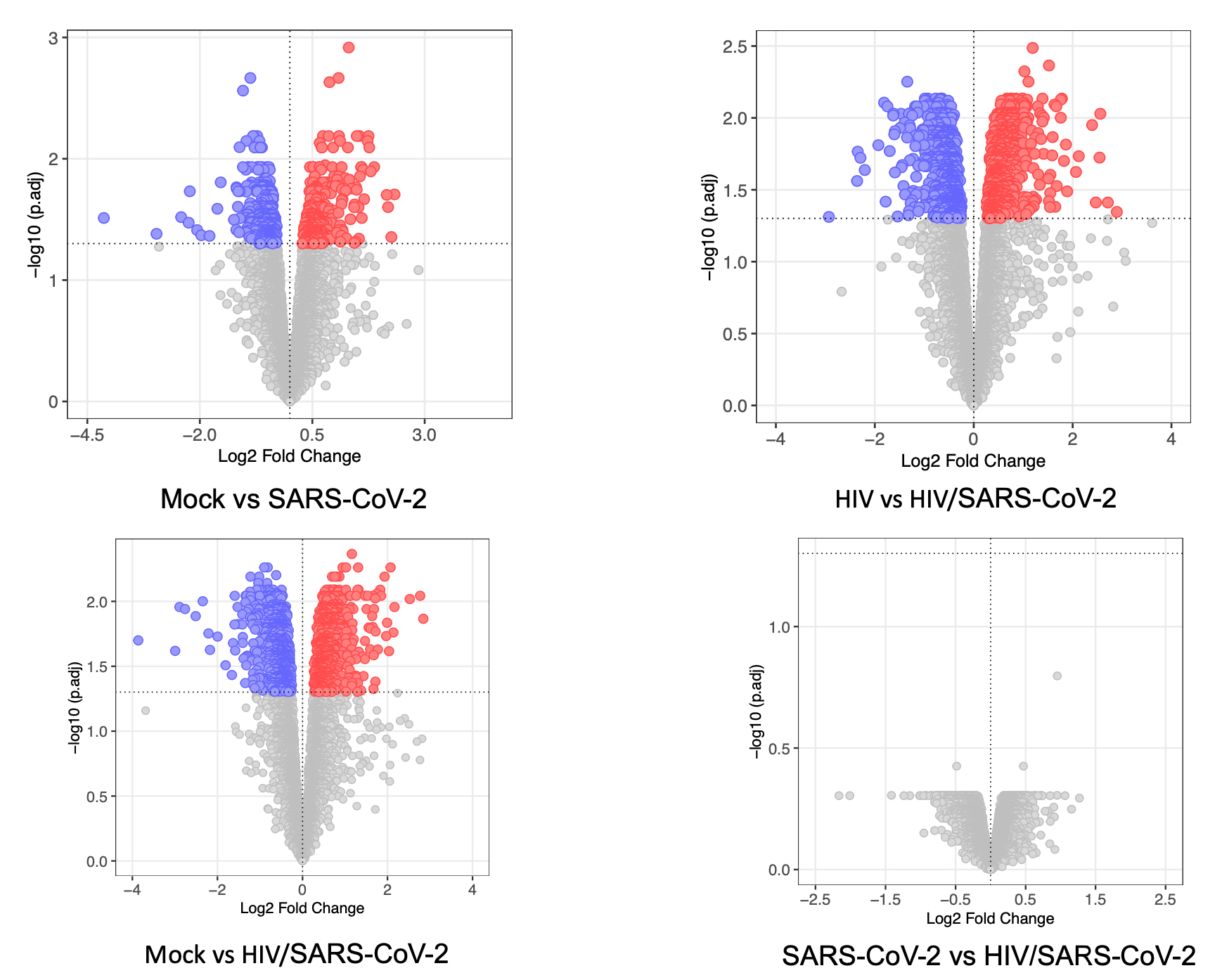
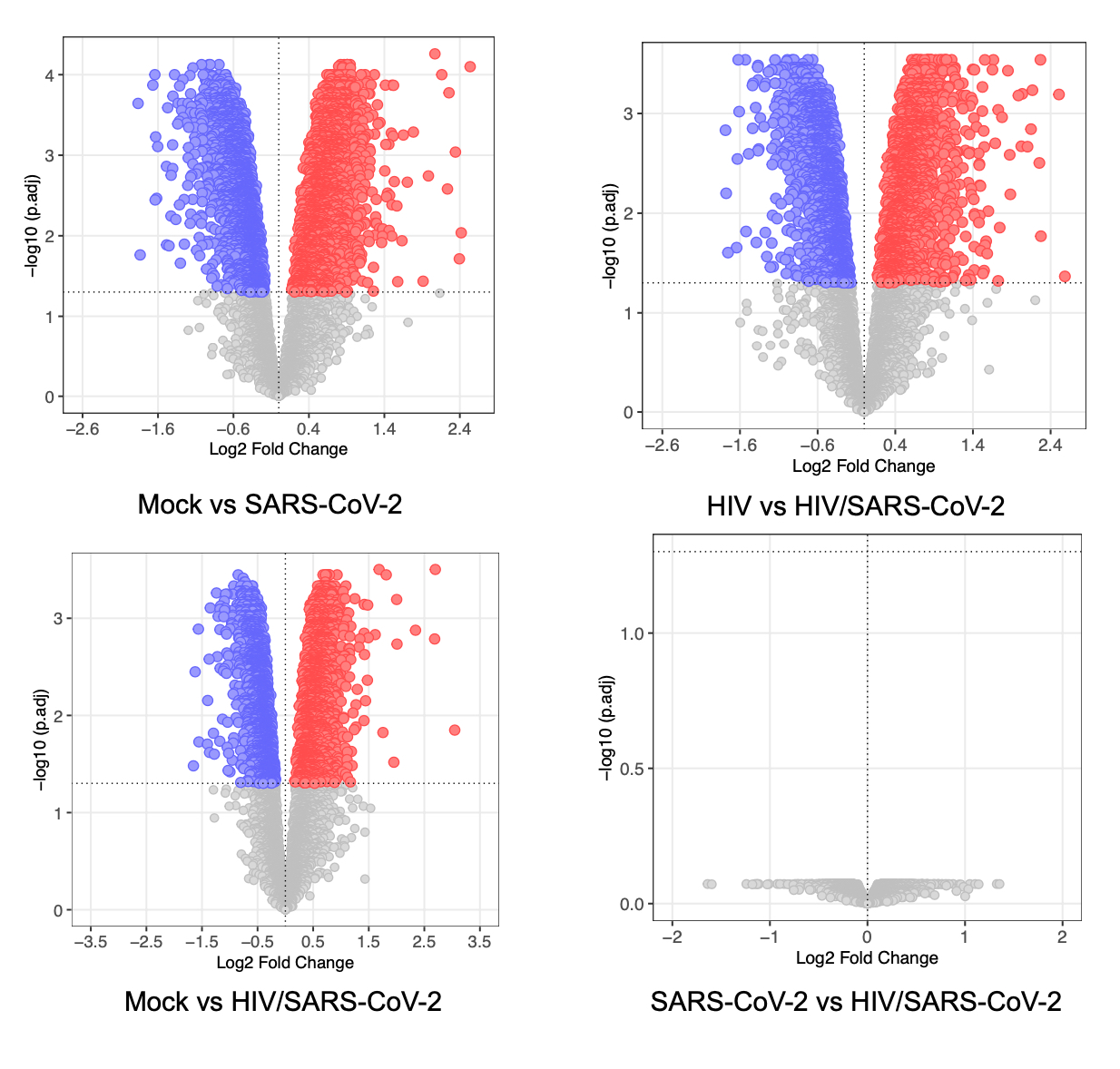
Many people after recovery forms of acute COVID-19 are suffering from PASC with cognitive impairments, and atrophy in the orbitofrontal and the para-hippocampal regions [45, 46]. These symptoms are similar to HAND observed in PWH [47]. Therefore, it is important to understand the long-term impact of COVID-19 in PWH. There is an ongoing argument on the mechanism of COVID-19-associated neuropathogenesis (i) Is it the result of direct invasion of SARS-CoV-2 in the brain or (ii) the consequence of systemic changes due to infection? Current data demonstrates a heterogeneous distribution of SARS-CoV-2 genomic RNA and viral proteins in brain even after recovery from active disease, and the underlying molecular mechanism remains unknown [42, 48–51]. Our findings indicate that even without establishing a productive infection, SARS-CoV-2 can induce a distinct proteomic signature in both healthy and HIV-infected astrocytes and pericytes.
In a Healthy brain, astrocytes perform various functions such as producing and releasing growth factors like NGF, BDNF, FGF-2, PDGF, GDNF, and TGFβ [52], maintaining BBB integrity, regulating cerebral blood flow, and water homeostasis, serving as an energy source for neurons, and maintaining the balance of glutamate-glutamine levels [53, 54]. However, during a pathogenic insult, in astrocytes undergo morphological, transcriptional, and functional changes, contributing to the development of neurodegenerative disorders [55]. Additionally, studies have shown that astrocytes can act as reservoirs for HIV reservoirs with restrictive replication and efficient cell-to-cell transmission capability [56, 57]. HIV infection induces activation of astrocytes, leading to the release of pro-inflammatory cytokine/chemokines and glutamate, causing excitotoxicity and neuronal death [58].
In our study, we observed differential modulation of energy metabolism pathways, such as Glycolysis/Gluconeogenesis and the Pentose phosphate pathway (PPP), in HIV/SARS-CoV-2 co-infected astrocytes. Dysregulation PPP has been linked to the development of neurodegenerative disorders line PD, AD, and ALS [59]. We also found enrichment of pathways related to propanoate metabolism, which is interconnected with beta-Alanine metabolism, and has been associated with AD development [60]. Additionally, we detect alteration in glycerophospholipid metabolism, that can impact anxiety-like behaviors, locomotor activity [61], and α-Synuclein mediated PD pathogenesis [62]. Astrocytes convert glutamate to glutamine, which helps maintain a glutamate/GABA balance in synaptic clefts and maintains the equilibrium in glutamatergic/GABAergic neurotransmission in CNS which is detrimental for its function [63]. HIV or its proteins like TAT/gp120 disrupt the Glu/GABA/Gln balance resulting in several neurodegenerative disorders [64]. Therefore, dysregulation in alanine-aspartate-glutamate metabolism in astrocytes of COVID-19 patients living with HIV may potentiate the development of neurocognitive disorders. Again, AD is associated with the dysregulation of several amino acid metabolisms [65] and we also detected enrichment of several amino acid metabolisms and degradation pathways in HIV/SARS-CoV-2 co-infected astrocytes. These findings suggest that PWH should be closely monitored for the development of AD-associated symptoms even after recovering from acute COVID-19 infection.
Pericytes are multifunctional mural cells surrounded by the basement membrane endothelial cells of the blood vessels throughout the body [66]. In the brain, pericytes remain embedded between brain microvascular endothelial cells, astrocytes, and neurons. They control BBB permeability, regulate angiogenesis and transmigration of systemic leukocytes in CNS, clear tissue debris, and Alzheimer's amyloid-β (Aβ) toxin from CNS [67]. Disruption of signaling pathways in pericytes can contribute CNS disorders like AD, and ALS [68–71]. Recent research suggests that pericytes also support productive HIV replication [72, 73]. Productive infection or exposure to HIV proteins results in the loss of pericytes from the BBB tight junction, which increases its permeability and facilitates the transmigration of systemic leukocytes in the brain parenchyma. This also leads to overexpression of pro-inflammatory mediators like IL-6, TNF-α, and IL-1β in pericytes, that triggers neuroinflammation [74, 75].
We identified specific cellular pathways that were differentially regulated in pericytes co-infected with HIV and SARS-CoV-2. These pathways include steroid hormone biosynthesis, thermogenesis, estrogen signaling, and ErbB signaling, which are all important for normal brain function [76]. Dysregulation in these pathways can have negative effects on neurogenesis, memory function, energy balance, and mental health [77]. Again, alteration in ErbB signaling in the brain, as observed in co-infected pericytes, is related to the development of depression [78]. This finding is also supported by the fact that many COVID-19 patients living with HIV develop depression after recovery from acute infection [79]. Mallat et al. describe the spike protein of SARS-CoV-2 damaging the vascular and immune regulatory functions of pericytes, resulting in vascular-mediated brain damage [21]. We also observed several immune-related pathways uniquely regulated in HIV/SARS-CoV-2 co-infected pericytes like Fc gamma receptor-mediated phagocytosis pathway, NK-cell mediated cytotoxicity pathways.
The limitation of the study includes the use of primary human astrocytes and pericytes. The human brain is a complex organ built with numerous cell types, which are interlinked with each other to form a sophisticated network that works in unison. The neurological consequence of HIV/SARS-CoV-2 may be a result of direct viral invasion to the CNS and neuronal injury, or indirect impact of systemic infection, involvement of a neuro-immune response, inflammation and immune activation, post-infection autoimmunity, and neurovascular injury, which is challenging to study using individual cell types [47, 80]. But no small animal model is available to study the impact of HIV/SARS-CoV-2 in the CNS, and the cerebral organoids used to study the SARS-CoV-2 associated neurological consequences are not susceptible to HIV infection. Other factors may impact the neurological outcomes, like whether the patients are on combined antiretroviral therapy or have complete plasma viral suppression, etc., which warrants the development of biologically relevant animal model to study in detail.
{kind=link}
{kind=link}
{kind=link}