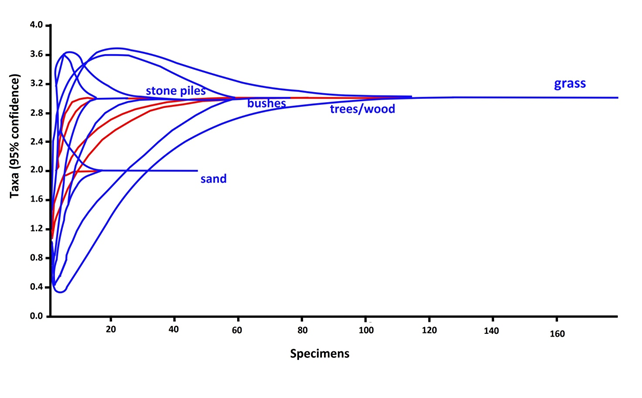
General community characteristics along the habitat gradient
Although not methodological consistent across habitat types, our study revealed that along the FSS gradient (i) there was a clearly higher species richness in the forest than in the savannah and suburbs, and that (ii) the three communities were quite taxonomically distinct. Thus, there was a “forest-specific” assemblage of species (including e.g. Varanus ornatus) and lowly diverse assemblages in savannahs (including e.g. Varanus exanthematicus) (Fig. 7) and suburbs, mostly characterized by ecological generalists with wide habitat tolerances. Both these diversity patterns were not unexpected (McKinney 2008). Concerning pattern (i), there is wide available literature showing that lizard communities are significantly more species-rich in tropical forests than in more open or degraded environments (Lewin et al. 2016), the same as generally observed in both vertebrates and invertebrates (e.g., Basset et al. 2008; Watling and Donnelly 2008). For instance, in the Philippines Supsup et al. (2020) found that higher species richness was found in secondary forests than in mixed-use agricultural areas. Since habitat heterogeneity is an important predictor in explaining lizard richness in Africa (Lewin et al. 2016), it is likely that the multiple microniches available in the hilly forests of the Togo hills may explain the much higher species richness that we observed therein in comparison with savannah and suburban habitats of the same study region.
Concerning pattern (ii), the fact that we observed habitat-specific assemblages of lizards at our study area mirrors observations from Brazilian Cerrado lizard communities (Nogueira et al. 2009). In the Brazilian Cerrado, in fact, habitat-specialists with patchy distributions in the forest-savannah-plantation mosaics create habitat-structured lizard assemblages with faunal overlap between forest and savannah being limited and forested versus open areas acting as mutual barriers to lizard distribution (Nogueira et al. 2009). In our study case, the taxonomical composition of the savannah assemblage was similar as that of the suburban habitat (although in the savannah we also observed Agama sankaranica and a few other species), thus suggesting that there is no clear barrier acting against lizard distribution between savannah and suburban areas in southern Togo.
The species richness reduction from forest to urban areas may also be interpreted as an effect of habitat loss on forest-specialists (Turner 1996; Lea et al. 2003; Edwards et al. 2013; Decena et al. 2020). Indeed, studies from tropical Africa have revealed that forest loss (by slash-and-burn shifting cultivations or by direct tree cutting) at the local scale produced a considerable reduction in the species richness of lizards (Akani et al. 2018). In a Niger Delta area, the number of sympatric lizards species was 9 in a unburnt forest patch, whereas it was just 2 in a nearby forest patch up to 12–16 months after fire (Akani et al. 2018). However, at the scale of southern Togo, the Dahomey Gap savannahs, albeit of likely anthropogenic origin, are there since few thousands years (XXXystemat; see Salzmann and Hoelzmann 2005). Thus, it is likely that the lower species richness of savannah lizard assemblages was due more to broad historical events and geographical/climatological reasons (Salzmann and Hoelzmann 2005) than to local-scale recent habitat loss.
Suburban lizards: analysis of community structure and niche characteristics
Our data revealed interspecific differences in both substratum type preferences and vertical spatial niche, but with no evidence of a mathematical structure consistent with a nonrandom niche partitioning pattern, and hence with a competitively assembled community structure. Instead, we detected a nonrandom “clustered” distribution of the various species along the available resource categories. Therefore, the main conclusion of our study is that species-specific preferences instead of community-driven mechanisms are more likely to explain the observed patterns at the study area.
Thus, we found no support for the hypothesis that lizards, as they can increase net energy gain from careful thermoregulation and hence selection of appropriate basking substrates, can compete for, and exclude potential competitors from, optimum basking sites (e.g.
Huey and Slatkin 1979). When interspecific competition cannot be advocated, the proximate external ecophysiological constraints are often considered the drivers of species-specific differences in niche characteristics (Barbault
1988; Barbault and Stearns
1991). In our case, thermal constraints pushing the selection of given substratum types for lizard basking (Huey and Slatkin 1979; Hailey
1982; Huey
1991) are unlikely to explain the observed patterns, as the study area has a tropical climate with ambient temperatures constantly ranging 27–32°C all throughout the year (
https://it.climate-data.org/XXXystem/togo/maritime-region/lome-764237/). Interspecific differences in substratum type uses due for foraging reasons are also unlikely as the various species have a relatively similar insectivorous diet: at the study area, the diet of
A. agama consisted mainly of Coleoptera adults and Formicoidea (Akani et al.
2013) and that of
T. quinquetaeniata of Coleoptera adults and larvae, Lepidoptera adults and larvae and Formicidae (Dendi et al.
2019). For
Hemidactylus sp. we do not have data for Lomé, but in a urban population from Port Harcourt (Nigeria) the diet was composed mainly by Lepidoptera adults and Formicoidea (Amadi et al.
2020a). In this latter species, however, the use of cement and elevated heights are likely linked to the need to exploit artificially lighted spots for optimal foraging over the insects that are attracted by night around electric bulbs (Amadi et al.
2020a). The same strategy was also employed by
A. agama in Nigeria (Amadi et al.
2021), but in Lomé we never observed any shifts from diurnal to nocturnal foraging activity in contrast with the Nigerian conspecifics (Amadi et al.
2020b,
2021), whereas dietary shifts have been observed in other tropical lizards from urban environments (Balakrishna et al.
2016). Thus, we suggest that antipredatory reasons may be behind the species-specific substratum type selection and the height of perches, given that predation risk is well known to be a crucial determinant of community structuring of tropical communities (e.g., Pianka
1973,
1986; Poulin et al.
2001; Olsson et al.
2005, Reyes-Olivares et al.
2020). We also noticed that the artificial substratum (cement) was predominantly used by both
A. agama and
Hemidactylus sp. In this regard, our data mirror other studies showing that lizards in urban areas use artificial substrates a large proportion of the time, and that these substrates tend to be broader than substrates in natural forest (Winchell et al.
2016; Amadi et al.
2020a,
2020b).
Intraspecific differences in the two species for which we collected such data (A. agama and T. quinquetaeniata) were minor, but A. agama males tended to select higher perches than females and juveniles. This positive selection for elevated perches is likely related to the complex hierarchic structure of the A. agama groups, with dominant males often XXXystematic adult females from higher height (Anibaldi et al. 1998).
General considerations on lizard communities in tropical cities
It is well known that urbanization, involving the conversion of natural habitats into human-modified ecosystems, usually reduce both the diversity and the abundance of indigenous animal communities, up to leading to the extinction of some species or even facilitating the establishment of non-indigenous communities in cities and towns (Hamer and McDonnell 2010). So, the general observations that we made (i.e. a reduced species richness in the suburban habitat of the FSS gradient) is not surprising. However, as mentioned above, our study also suggests that the lizard species used the available resources in a clustered way within the suburban habitat in Lomé. Whereas many studies analyzed reptile assemblages in secondary forests and altered habitats (e.g., Luya et al. 2008), the data available in the literature with regard to communities of lizards (or other reptiles) in tropical cities are relatively few and based on different methodologies from ours, and so comparisons with our study are not straightforward. However, in a snake community inhabiting a Brazilian city, Franca and Franca (2019) observed a clustered distribution for two species and dispersed distributions for eight species, and in a Indian city, Janiawati et al. (2016) demonstrated that the various reptilian species were clustered around water sources and vegetation cover patches. In fact, in urban fragments embedded in an “artificial matrix” hostile to the movement of individuals, most species tend to be concentrated into suitable microhabitats (and using clustered types of substratum) as extinction rates (stochastic or anthropogenic) are not compensated by colonization rates (Laurance et al. 2006a, 2006b; Salomão et al. 2019).Thus, we may conclude that lizard communities in tropical urban areas are (i) less species-rich than in the surrounding more natural habitats, (ii) usually clustered into specific habitat/substratum types (often artificial ones), and (iii) not assembled through competitive interactions. Further studies should verify the validity of these conclusions.
{kind=link}