Metabolic stress aggravates PTOA development
Diet-induced metabolic stress was observed in the HFD group, as demonstrated by the metabolic profile of the HFD animals at the end of the study (Table 2). Metabolic stress was already present before surgery: body weight, cholesterol levels, and HOMA-IR were significantly increased in HFD mice compared with LFD controls after 9 weeks (data not shown). Body composition analysis revealed that the increase in body weight was completely attributed to an increase in body fat mass.
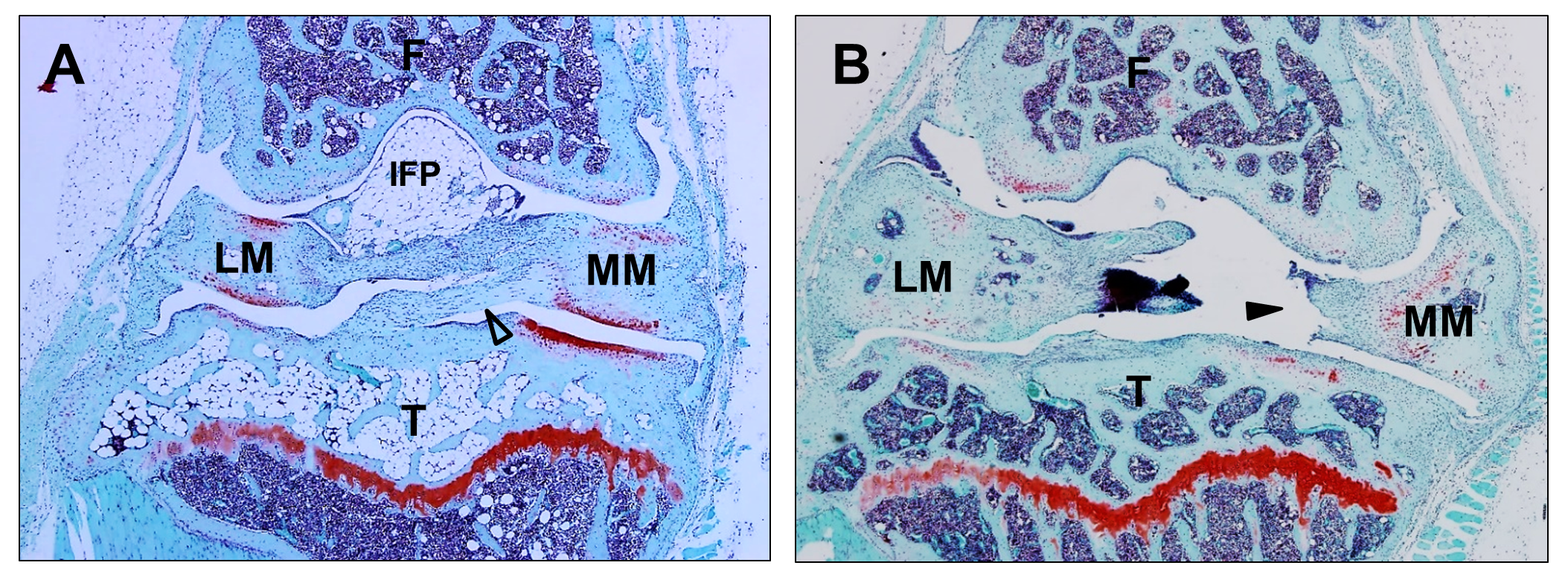
Articular cartilage damage was assessed by histological examination to evaluate the effect of HFD on DMM-induced cartilage damage. In the sham-operated contralateral knee joints hardly any cartilage damage was observed (Figure 1). DMM surgery on the other hand induced clear OA development in both groups, with diet-induced spatial differences in cartilage damage (Figure 1). Cartilage damage was significantly aggravated in HFD mice compared with controls, with median [interquartile range] OA scores of 19.4 [15.1-20.9] in HFD versus 13.8 [8.0-20.0] in LFD mice (p=0.049, Figure 2A). This difference could mainly be attributed to increased damage in the lateral tibiofemoral compartments (Figure 2B). HFD mice showed advanced cartilage erosion in both lateral compartments, extending to 25-50% of the articular surface (8.0 [4.9-10.0]; p=0.01), while LFD mice showed increased fibrillation, fissures and occasional cartilage loss <25% (3.8 [3.0-8.0]) (Figure 1). The medial knee compartments, typically more affected in the DMM model, were almost completely devoid of cartilage and reached close to the maximal score of 12 in both the HFD and LFD groups (Figure 2C). Therefore, even if potential diet-induced aggravation had occurred, this could not be inferred from the medial side as these were nearing maximal scores (HFD 11.0 [10.6-11.8]; LFD 10.0 [5.0-12.0]; p = 0.109).
Synovial inflammation was significantly increased and more severe in HFD mice (7 out of 8 mice; 6.5 [5.3-8.5]) compared with LFD mice (4 out of 7 mice; 1.0 [0.0-6.0]; p=0.04, Figure 2D). Sham-operated contralateral knees showed no synovial thickening. Increased osteophyte formation was observed in the HFD group compared with the LFD controls (Figure 2E). Sham-operated contralateral knee joints showed hardly any osteophyte formation.
Systemic changes in eicosanoid levels
Eicosanoids can exert both pro- and anti-inflammatory functions and have been implicated in the pathogenesis of a variety of immunometabolic disorders, like obesity13 [17]. Eicosanoids are formed from omega-3 (n-3) or n-6 polyunsaturated fatty acid (PUFA) precursors via the cyclooxygenase (COX), lipoxygenase (LOX), and cytochrome P450 pathways (CYP450, Figure 3)[22]. At large, n-6 PUFA-derived eicosanoids exhibit more pro-inflammatory and n-3 PUFA-derived eicosanoids more anti-inflammatory activity. Although a useful framework, this simplistic dichotomy does not cover the complexity of the physiological effects of eicosanoids in vivo[22].
Because of plasma volume limitations and the volume demands of the assay, we were able to reliably detect the levels of 24 eicosanoids. Overall, minor differences were observed between the diet groups. HFD induced a significant increase in arachidonic acid (AA) levels, a n-6 PUFA precursor, with 947 [765-1257] in LFD versus 1538 [1230-1935] in HFD mice (p<0.01) at 14 weeks. To find out how this increase affected total eicosanoid metabolism, we evaluated the changes in eicosanoid profiles of the mice before and after DMM surgery. Pathway analysis revealed that the increase in AA levels mainly translated into increased hydroxylation into the fatty acid diols 5,6-dihydroxyeicosatrienoic acid (DHET) and 8,9-DHET (Figure 3). In addition, 5-HETE and 13-OxoODE levels were increased in HFD mice compared with LFD mice. Similar plasma eicosanoid profiles were observed between 9 and 14 weeks (data not shown).
Activation of circulating monocyte subpopulations
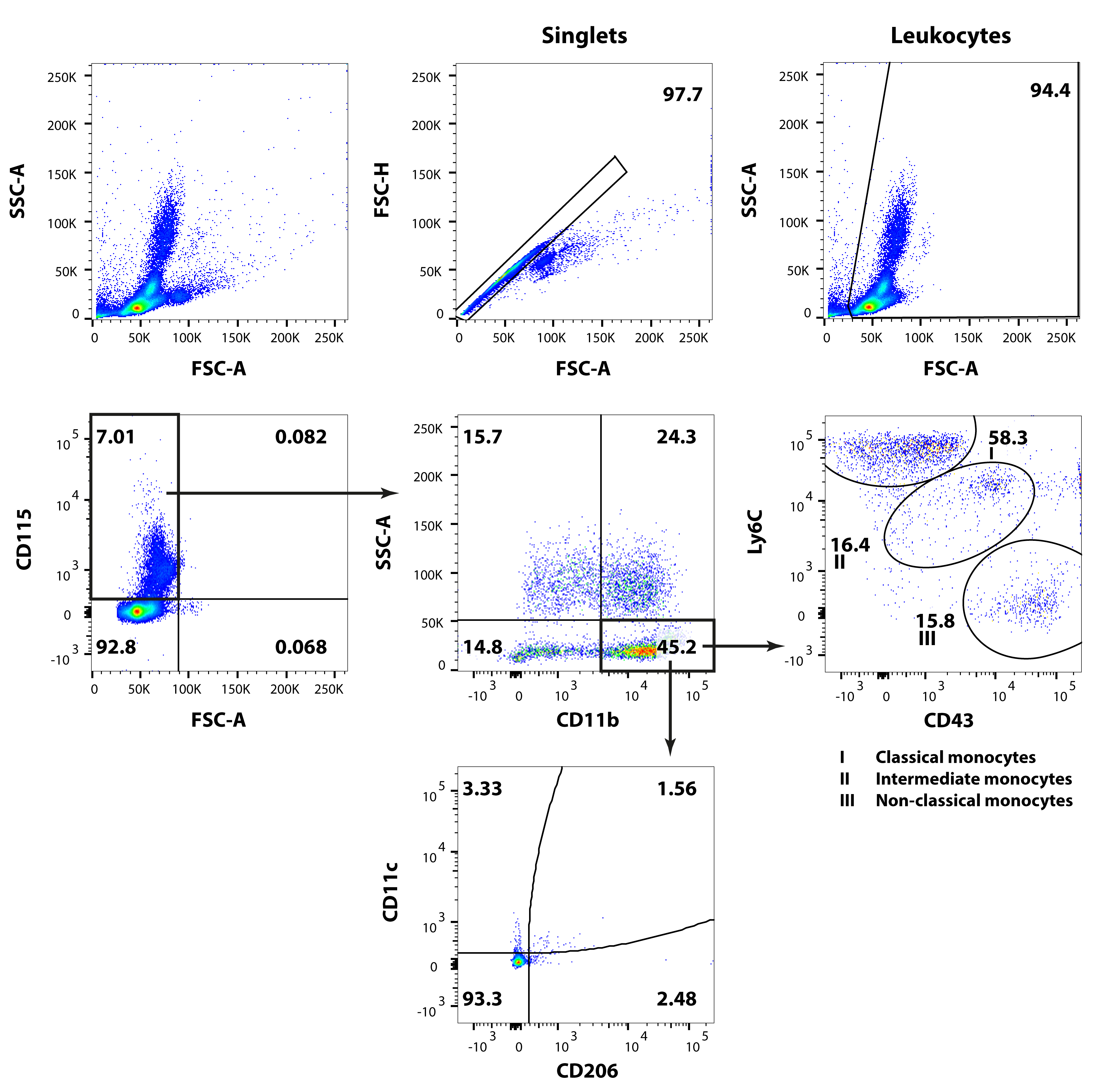
Monocytes are short-lived mononuclear phagocytes that constitute ∼10% of peripheral leukocytes in humans and ∼4% in mice[23]. Current nomenclature divides murine monocytes into three subpopulations based on their surface expression of CD43 and Ly6C receptors[24]: 1) CD43+Ly6Chi classical monocytes; 2) CD43++Ly6Cint intermediate monocytes; 3) CD43++Ly6Clow non-classical monocytes. The three subpopulations share a developmental relationship (from classical via intermediate to non-classical) and are phenotypically and functionally different.
Monocyte subpopulation percentages showed subtle differences between the LFD and HFD groups over time (Figure 4A). The most pronounced effect of long-term HFD was an increase in circulating CD43++Ly6Cint intermediate monocytes at t=17 weeks, at the expense of the CD43++Ly6Clow non-classical subpopulation. This increase showed significance within the total CD115+CD11b+SSC-Alow monocyte fraction (p = 0.044, data not shown), but due to interindividual variability did not reach significance within the total leukocyte fraction (p = 0.107, Figure 4B).
The increase in intermediate monocytes concurred with a significantly increased expression of the monocyte activation marker CD11c in HFD mice compared with LFD controls (0.8 [0.7-1.4] versus 3.0 [1.6-3.9] at 9 weeks and 3.0 [1.9-3.4] versus 6.6 [4.6-7.7] at 17 weeks, respectively; Figure 4C). CD11c contributes to monocyte arrest on endothelial cells and, as such, is upregulated by activated monocytes during chemotaxis. Both high-fat diet and knee injury contributed to the increased expression of CD11c, respectively, as: 1) HFD mice showed significantly more CD11c expression compared with LFD controls at 9 and 17 weeks (p<0.01) and 2) LFD mice showed increased CD11c expression over time (p=0.03). Backgating (i.e. highlighting the final gated population within the population of its ancestors) of the CD11c+ cells into the total CD115+CD11b+SSC-Alow monocyte fraction revealed that this integrin was predominantly upregulated by the CD43++Ly6Clow non-classical monocytes (Figure 4D).
High-fat feeding induced no macrophage polarization
As activated monocytes may differentiate into macrophages upon tissue entry, the effects of the HFD on this immune cell type were investigated locally by evaluating the surface area percentage positivity of M1 (iNOS) and M2 (CD206) macrophage markers in the synovial lining of DMM-operated right knee joints.
Synovial thickening was increased in knee joints of HFD mice compared with LFD mice (Figure 5A). The ratio of the surface area percentages between M1 marker iNOS and M2 marker CD206 showed an increase in the HFD mice that did not reach significance (LFD 0.54 [0.39-1.34] versus HFD 1.28 [0.62-1.66]; p=0.12; Figure 5B). Pan-macrophage marker F4/80 positivity was predominantly observed in the upper synovial layer and showed a non-significant increase on HFD (0.94 [0.20-2.78] versus 2.49 [1.61-5.25]; Figure 5C). The subset-specific markers CD206 and iNOS were upregulated in the HFD group compared with LFD group as well, but none of the markers reached a significant difference between both groups. This was also observed within the F4/80+ macrophage fraction, where the colocalization surface area percentages of CD206 and iNOS positivity were similar between diet groups (Figure 5D).
Correlation analyses revealed moderate to high positive associations for the total F4/80-stained surface area of the synovial lining with lateral OA degeneration and synovitis scores (Table S2). Both OA scores significantly correlated to the colocalization of the total area positivity of the three macrophage markers as well. Osteophyte scores showed no correlation with any of the macrophage stains. Within the F4/80+ fraction, neither subset marker associated with the evaluated OA features.
{kind=link}
{kind=link}