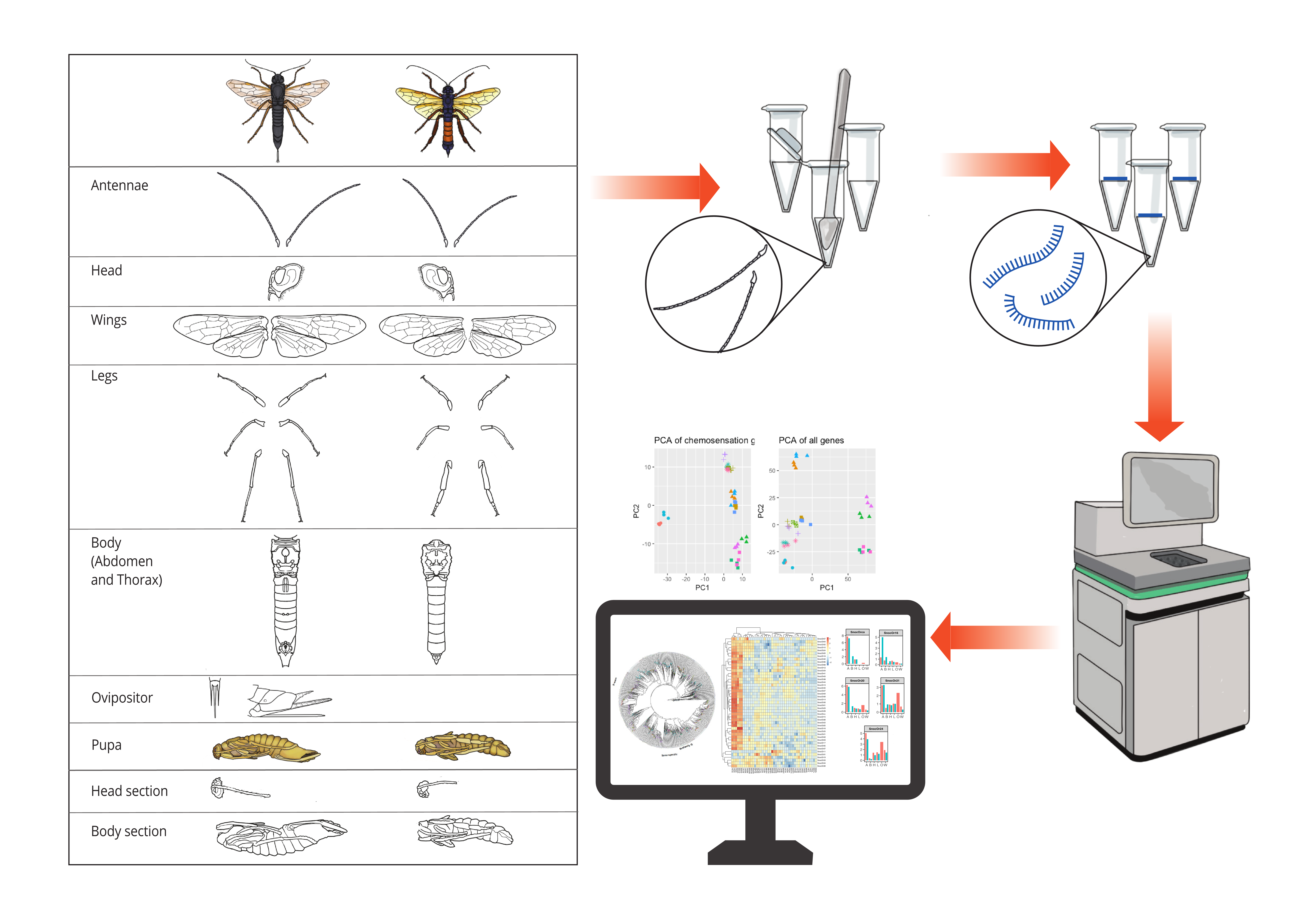
Comprehensive sawfly chemosensation gene repertoire
One hundred and twenty-nine chemosensory genes were identified in the genome of the invasive woodwasp, Sirex noctilio, including 45 ORs, 12 GRs, 48 IRs, 14 OBPs, 9 CSPs and one SNMP. All identified receptor sequences were predicted to contain chemoreceptor functional domains. Binding proteins (OBPs and CSPs) contained binding protein domains and signal peptides.
A single CO2 receptor ortholog (SnocGR11) was identified for S. noctilio. Two CO2 chemoreceptor orthologs were identified in C. cinctus [1]. The presence of these orthologs in the Symphyta is in contrast to species of the Apocrita (e.g., N. vitripennis, A. mellifera, and B. terrestris) that appear to have lost this receptor, as none have a CO2 receptor ortholog [44]. Insects detect CO2 for a variety of reasons ranging from avoidance of harmful levels of CO2 [45], host location in mosquitoes, flower quality assessment in Lepidoptera, brood placement and maintenance in social insect colonies [46], as well as oviposition in phytophagous insects [47]. The CO2 GRs of C. cinctus were expressed in head tissues, however, low expression levels of the S. noctilio orthologue were observed in this study.
SnocGR1 and SnocGR2 are orthologous to AmelGR10, a broadly tuned amino acid receptor with high expression in the fat body of A. mellifera [48]. Interestingly, homologs of AmelGR10 are conserved amongst Hymenoptera, but not found in non-eusocial species [48]. Insect fat bodies are responsible for energy metabolism and nutrient storage and undergo reorganisation during metamorphosis [49]. Expression profiling of AmelGR10 revealed differential expression across developmental stages [50]. In S. noctilio, we found that SnocGR1 and SnocGR2 had overall low expression levels across all tissues with the highest levels in pupae, suggesting that they might be involved in growth and development processes.
Global gene expression patterns were distinct between developmental stages rather than tissues. However, when focussing on chemosensation genes, antennae clustered separately from all other conditions (tissues and developmental stages). This highlights the importance of antennae in chemosensation, and particularly olfaction. Compared to other tissues, antennae displayed the greatest number of differentially expressed chemosensory genes. Of these SnocOBP9, 10, 14, SnocCSP7 and SnocGR9 appear to be of particularly interesting olfaction gene targets based on the antennae-biased expression profiles of these genes.
Of the 6 gene families studied, ORs displayed the greatest expression bias in antennal tissues. We observed that the co-receptor, SnocORco, was expressed in all tissues and at higher levels than any of the other chemoreceptor genes. The antenna-biased expression of ORs and expression levels observed for ORCO, are consistent with what has been reported in other studies. ORCO improves the responsiveness and sensitivity of ORs and enables accurate detection of chemical stimuli which could aid in a variety of ecologically important chemosensory functions [11], such as oviposition-site selection and mate location.
Dimorphism of Sirex noctilio chemosensation: Tissue and sex differences
The comprehensive investigation of the phylogenetic relationship and expression patterns of chemosensation genes in this study led to the discovery of several intriguing gene targets for further investigation of pathways underlying ecological specialisation and success of this invasive woodwasp. For example, two odorant binding proteins (SnocOBP2 and SnocOBP5) are shown to be putatively involved in oviposition site selection. Another example is SnocIR09 that is uniquely expressed in wings and might be involved in mate or host location. In the rest of the discussion, however, we have focused on genes possibly involved in olfaction.
The results of phylogenetic and gene expression analyses suggest the potential involvement of certain ORs in sex-specific behaviour of S. noctilio. SnocOR6, 9, 22, 23, 24, 28 and 39 were significantly upregulated in female antennae compared to male antennae. These genes form part of a group of thirteen S. noctilio ORs (SnocOR9, 13, 14, 23, 25, 28, 29, 30, 38, 39, 40, 41, 44) which are possibly the result of species-specific expansion and form a unique phylogenetic clade within the OR gene family. Some of these S. noctilio genes (SnocOR9, 14, 28, 29, 31, 40) were also found to be orthologous to ORs from the bark beetle, I. typographus. It is predicted that receptors encoded by these bark beetle genes interact with terpenoids from host trees and symbiotic fungi [42]. This is a potentially important discovery since the S. noctilio orthologs might have a similar function responding to volatiles of host trees and symbiotic fungi. Females are expected to express these receptors at higher levels than males since they need to find suitable hosts for oviposition.
SnocOR24 was significantly upregulated in female antennae and biased expression was also observed in ovipositor tissues. SnocOR24 is at the base of a large clade that contains the A. mellifera gene, AmelOR11. AmelOR11 is the honeybee OR responsible for the reception of the queen pheromone component, 9-oxo-2-decenoic acid, and is one of the few olfactory receptors that have been deorphanised [51, 52]. The orthology to the honeybee pheromone receptor, as well as its female-biased expression patterns, indicates that SnocOR24 may be involved in S. noctilio female sexual behaviour.
SnocOR20 and SnocOR21 exhibited high expression levels in antennae and SnocOr20 was significantly upregulated in antennae compared to other tissues. These two genes are orthologues of the A. mellifera gene, AmelOR151, which encodes a receptor that responds to the floral terpene, linalool [53]. Linalool has several reported biological functions, including anti-microbial, anti-oxidant, larvicidal, insecticidal, pollinator attractant and repellent activities [54, 55]. Amylostereum areolatum, the symbiotic fungus of S. noctilio, is known to produce a four-compound blend which includes linalool to which female wasps are attracted [56]. In C. cinctus, the wheat stem sawfly, behavioural studies showed that host selection by ovipositing females is influenced by the host volatiles (Z)-3-hexenyl acetate and linalool [57]. Achhami et al. [57] suggested that high levels of linalool in the host plant may deter oviposition by C. cinctus females. Thus, SnocOR20 and SnocOR21 might play a role in recognition of potentially ecologically important symbiont or host volatiles.
The expression profile of SnocOBP2 in the female ovipositor suggests chemosensory functions possibly related to oviposition-site selection. Female woodwasps use their ovipositors to probe a host tree to determine whether it is a suitable site for egg development. The ovipositor of S. noctilio has also been shown to have electrophysiological activity capable of pine volatile and ethanol detection [40]. Furthermore, Faal et al. [56] also showed that S. noctilio females are attracted to volatiles emitted from A. areolatum, thus SnocOBP2 could also be involved in detection of fungal volatiles from fungal infections from previous wasp attacks.
Normalised gene count data showed that male leg tissues clustered separately from wings, ovipositor, and female leg tissues. Adult S. noctilio are sexually dimorphic and males have larger hind legs than females. Hind legs of S. noctilio have been hypothesized to be the site of synthesis and/or storage of a putative male-produced pheromone blend that may act as an aggregation-pheromone [38]. This pheromone could mediate lek formation in male S. noctilio. Therefore, genes such as SnocOBP3 and SnocSNMP1 are of interest due to their enhanced expression in male leg tissues.
SnocOBP3 was adult male leg-biased suggesting possible involvement in male-specific behaviours. Previous transcriptome studies suggested leg-biased OBPs may mediate host-seeking behaviours [58–60]. However, unlike SnocOBP3, OBPs from these studies were not sex biased. The male leg-biased expression of SnocOBP3 could imply a role in pheromone detection and/or the release and storing of chemical compounds [4]. Female-biased cuticular hydrocarbons (CHCs), 4-allyl-2,6-dimethoxyphenol (4-DMP), produced by Bactrocera dorsalis fruit fly females act as a close-range sex pheromone that is attractive to males. No response was elicited toward 4-DMP in male B. dorsalis antennae during electroantennography (EAG) [61]. However, an electrophysiological response was detected in male mid-legs during EAG with 4-DMP [62]. Like S. noctilio, legs of B. dorsalis males are in close contact to female bodies during copulation. Exposure to 4-DMP led to downregulation of BdorOBP2, an OBP with strong binding affinity to 4-DMP and therefore, a candidate transporter of 4-DMP [62]. Reduced transcript abundance led to reduced behavioural response in male flies. The downregulation of binding proteins has been hypothesised to be a mechanism to regulate overstimulation of the chemosensory system [63]. It would be worth studying the possible involvement of SnocOBP3 in CHC perception in male leg tissues during copulation.
Antennae and male leg biased expression of SnocSNMP1 is consistent with the hypothesis that SNMP1 is required along with pheromone-detecting ORs to recognize lipophilic pheromones [64]. The expression profile of SnocSNMP1 was also observed for A. cerana cerana where SNMP1 is highly expressed in antennae and legs [65]. The study in A. cerana cerana suggested that the expression of SNMP1 in legs may indicate involvement in various sensory processes. Demonstration of S. noctilio male hind legs as the site of synthesis and/or storage of a putative aggregation-pheromone [38] may implicate the expression of SnocSNMP1 in male legs in pheromone detection. Cumulatively the phylogenetic and gene expression data indicate that this gene may play a role in sex-specific behaviours of S. noctilio.
SnocOR16 was expressed at significantly higher levels in male antennae, a result also observed by Guo et al. [41]. This receptor clustered at the base of the 9-exon subfamily which is known to be involved in the reception of CHCs and has been implicated in social interactions within the Hymenoptera [43, 66–68]. Cuticular hydrocarbons cover the insect cuticle and protect against desiccation and mediate social behaviours, ranging from courtship and aggregation to colony organisation and conspecific communication especially in eusocial insects [43, 66, 69–72]. Ants discriminate between CHCs without contact between individuals implicating CHCs as potential olfactory cues detectable by ORs housed in antennae [73]. Sirex noctilio females produce CHCs (monoalkenes) on their cuticles that induce male copulation [37] and it could also play a role in other recently reported mating behaviours. For example, Queffelec et al. [74] found that female attractiveness decreases with age and that male S. noctilio wasps exhibit potential mate choice. Queffelec et al. [74] proposed that production of CHCs by female wasps could decrease with age making older females less attractive and subsequently have fewer matings. Deorphanisation studies between SnocOR16 with CHC ligands could elucidate the potential role of this receptor in the sexual behavioural ecology of S. noctilio.
{kind=link}
{kind=link}