Microplastics (MPs) are ubiquitous in our environment due to human activities. Recent studies have increasingly confirmed the presence of MPs in the human body, raising concerns about their potential hazards. However, the impact of PS-MPs on obesity of human remains unclear. Considering that HFD effectively induces obesity, and HFD combined with other factors (e.g., Azithromycin) may aggravate obesity [52], in this study, we set out to examine the detrimental impact of PS-MPs when administered through drinking water in a HFD-induced obesity model. Our study presented compelling evidence that PS exacerbated HFD-induced obesity and promoted weight gain, which was associated with impaired glucose and lipid metabolism. As a considerable proportion of ingested MPs traversed the gastrointestinal tract following oral intake [31], it was hypothesized that the gut microbiota and epithelial barrier may represent novel targets for PS-MPs toxicity, particularly in obese populations. By utilizing a mouse model of obesity induced by a HFD, and employing ABX treatment and FMT, this study has furnished compelling evidence that PS treatment induced inflammation and obesity by altering the composition of the gut microbiota and compromising the integrity of the intestinal barrier. To our best knowledge, this is the first study to explore the obesity-enhancing effects of the combination of a HFD and PS, as well as to evaluate the role of PS-induced alterations of gut microbiota in HFD-induced obesity. Our findings suggested that PS might exacerbate Western-diet induced obesity and related metabolic disorders in humans, and the gut microbiota is a crucial target for the obesity-promoting effects of PS-MPs.
The daily intake of MPs is still uncertain, despite organisms being constantly exposed to them. This is particularly true for humans, as the pathways of MPs contact are diverse and complex, leading to varying assessments of body intake. The dose selection for PS-MPs took into account both their toxicological and environmental significance. PS-MPs with a yield of 5 µm and a toxic effect were chosen as exposure materials, based on previous studies on male intestinal toxicity, with a range of 5 µg/d to 1 mg/d [30, 53–55]. In terms of environmental significance, on average, humans could potentially ingest 0.1-5 g of MPs per week globally [56]. Based on the weight of the mice, which was approximately 20 g, their daily intake of PS was calculated to range between 37.14 µg/d and 1.86 mg/d. It was estimated that each control mouse consumed 3.6 mL of water per day, resulting in a PS intake of 36 µg/d per mouse [32]. This amount is comparable to the minimum intake for humans. The exposed doses of mice were exposed daily to PS through their drinking water, simulating the major route of PS exposure in mammals [57, 58]. Therefore, the selected doses were of environmental and toxicological significance.
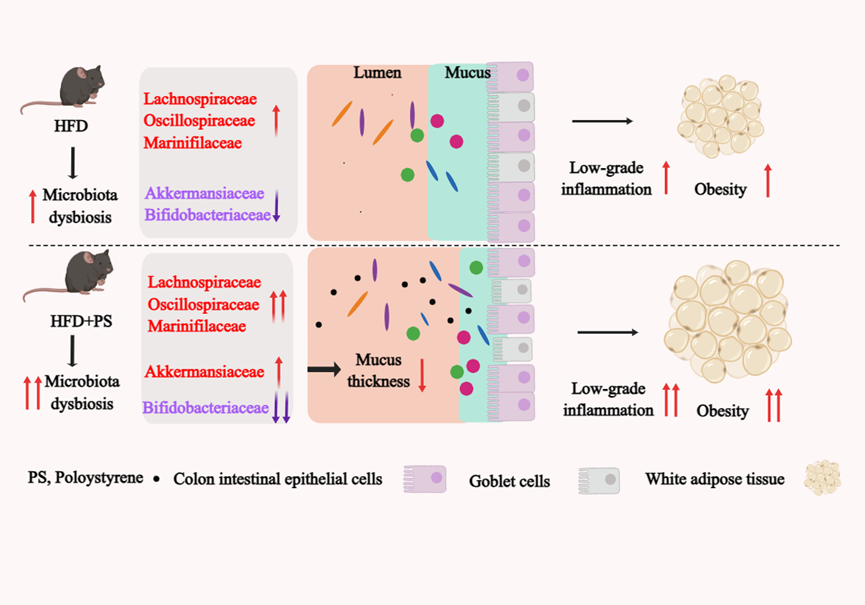
Recently, it has been discovered that MPs have the ability to disrupt glucose and lipid metabolism in rodents [59], however, the role of PS in obesity populations is not yet well understood. Our study first confirmed that PS exacerbated HFD-induced obesity. It is noteworthy that the lipid accumulation in the liver and white adipose tissue caused by PS was more pronounced in HFD-fed mice than in those fed a ND, which suggested that susceptible populations might be more affected by the impact of PS. This suggested that PS-induced obesity was related to diets. This could be attributed to the fact that, in addition to PS consumption, HFD also caused mucosal damage and low-grade inflammation [60]. At the molecular level, the process of adipocyte differentiation is controlled by multiple transcription factors, which facilitate the expression of adipogenic genes and support cell differentiation. Notably, recent research has highlighted the significant role of PPAR-γ and C/EBP-α in regulating this process. [61]. PGC-1α plays a vital role in maintaining energy balance by regulating the transcription of genes responsible for fatty acid oxidation [62]. In our study, PS administration up-regulated adipogenic genes such as PPAR-γ and C/EBP-α, and down-regulated lipolysis genes PGC-1α in the liver of mice fed a HFD. It was suggested that the pro-obesity activity of PS might be mediated by up regulating the expression of transcription regulator genes of lipogenesis and down regulating the expression of transcription regulator genes of lipolysis, which was further corroborated by fat accumulation and the elevation of TG and TC levels in the liver. It is well known that overweight and obesity are associated with significant changes in metabolism, especially glucose and lipid metabolism [63]. We consistently found that glucose tolerance was severely impaired in HFD + PS group, and serum levels of TC, LDL-C and LDL/HDL-C ratio were significantly elevated. Together, our results indicated that alterations in glucose and lipid metabolism played a significant role in the development of PS-induced obesity. Recent studies have revealed that the development of obesity in mice exposed to PS can be effectively dose-controlled under ND conditions [64]. Specifically, relatively low doses of PS have been found to trigger obesity while higher doses do not. In line with these observations, we report that administering 10 mg/L PS in drinking water did not induce obesity in mice on a ND. Intriguingly, and in contrast to the above findings, we observed that 10 mg/L PS exposure promoted fat accumulation in mice fed a HFD, suggesting that PS-induced obesity was highly influenced by diets as well as dosage of PS. Previous studies have reported that oral intake of 0.45–0.53 µm PS-COOH beads or a combination of PS particles with varying particle sizes (5, 50, 100, and 200 µm) led to a reduction weight in HFD-fed mice [49, 65]. This implies that PS with distinct particle sizes and properties may have dissimilar effects on body weight. In the following phase, we will investigate the impact of PS and other microplastic particles on obesity in mice under varying dietary conditions and dosage levels. Our findings demonstrated that exposure to PS at levels below those typically encountered by humans worsened HFD-induced obesity, potentially elucidating the link between increased plastic production and the growing global prevalence of obesity.
Recent studies have shown that PS-induced obesity is related to gut microbiota [32, 64]. Previous research has revealed that obese mice have different intestinal compartments compared to their normal counterparts, with a higher proportion of Firmicutes phylum [66, 67]. Additionally, it has been suggested that Firmicutes phylum may be implicated in obesity, as a larger population of these microorganisms can effectively convert food to energy at accelerated rates [68]. The study found that PS increased the relative abundance of the class Clostridia (phylum Firmicutes), including family Lachnospiraceae, and Oscillospiraceae in the HFD + PS group. Interestingly, PS was also able to reduce the F:B ratio, which is considered a key marker of diet-induced obesity [9, 10]. These findings suggest that PS can modify the population of certain bacteria within the Firmicutes phylum, leading to obesity independent of the F:B ratio.
Therefore, the relative abundance alterations of certain family levels within the Firmicutes bacteria was a significant factor in PS-induced obesity. Previous studies have reported an relative abundance increase of family Lachnospiraceae in both mice and humans with obesity and diabetes [69, 70]. Mechanistic research has shown that Fusimonas intestini, a commensal species of Lachnospiraceae, contributes to diet-induced obesity by producing excessive long-chain fatty acids [71]. In our study, Lachnospiraceae was the predominant family of Firmicutes, comprising over 40% of the total population. The overproduction of long-chain fatty acids could potentially be linked to PS-induced obesity. Family Oscillospiraceae was reported to be increased in HFD-fed mice. This family exhibited a positive correlation with obesity indexes, blood parameters, bacterial translocation parameters, proinflammatory cytokine, TLR4/NF-κB pathways, energy metabolism, and colonic permeability [45, 72]. Consistent with these observations, PS treatment significantly enhanced the percentage of family Oscillospiraceae in the gut microbiota of the HFD + PS group. In addition to alterations in bacterial abundance within the Firmicutes phylum, there is a remarkable relative abundance increase in family Marinifilaceae in HFD + PS group, which were significantly associated with visceral fat and body fat percentage [73]. Correlation analysis also proved that the relative abundance of family Lachnospiraceae, Oscillospiraceae and Marinifilaceae were significantly positively correlated with obesity related indicators.
In particular, the relative abundance of genus Akkermansia significantly increased in the HFD + PS treatment group. Recent research has highlighted the significance of genus Akkermansia as an essential functional bacterium that regulates human immunity and metabolism in the intestine, significantly linked with human health and diseases [74, 75]. Normally, genus Akkermansia colonized the outer mucous layer of the gastrointestinal tract to maintain the dynamic balance of mucin production, thereby upholding the stability of the mucus layer [76]. Furthermore, genus Akkermansia could significantly improve metabolic endotoxemia, insulin resistance, fat accumulation and fasting hyperglycemia in obese mice induced by HFD [77]. However, genus Akkermansia's role in intestinal diseases remains controversial. Some studies have found that genus Akkermansia could improve obesity and obesity-associated metabolic syndrome in mice [77, 78]. In contrast, clinical studies have revealed a high frequency of genus Akkermansia in the fecal matter of infants with eczema. Moreover, it has been established that genus Akkermansia impairs gut barrier integrity and elevates the risk for eczema [79]. Recently, a study demonstrated that a high-sugar diet could augment the relative abundance of genus Akkermansia in mice. This increased abundance resulted in the destruction of intestinal barrier function, leading to colitis via stimulating the activity of mucolytic enzymes within the microbiota [80]. In this study, PS was observed to significantly enhance the relative abundance of genus Akkermansia, while decreasing the thickness of the mucus layer in the colon tissue of HFD-fed mice, which potentially contributed to worsening obesity.
Numerous reports have indicated that the onset of obesity induced by dietary factors is linked to changes in gut microbiota, characterized by a reduction in beneficial microbial genera, which ultimately contributes to obesity and metabolic syndrome [81–83]. Consistently, our study also revealed a reduction in the abundance of beneficial bacteria, including family Atopobiaceae and genus Bifidobacterium, as well as genus Parabacteroides. The function of family Atopobiaceae is not yet fully understood; however, previous studies have reported a significant decrease in the abundance of family Atopobiaceae in HFD-induced diabetic mice. [84]. Consistent with the findings of this study, the relative abundance of family Atopobiaceae was observed to be negatively correlated with the obesity index induced by PS, indicating a potential role of family Atopobiaceae in anti-obesity. Previously, the Bifidobacterium spp. had been shown to reduce obesity [85], which was negatively related to intestinal permeability, metabolic endotoxemia, and low-grade inflammation [86, 87]. Similarly, this study found that PS promoted a significant decrease in the proportion of genus Bifidobacterium in HFD-fed mice, which was a negative correlation with obesity-related indicators. Except for genus Bifidobacterium, a study revealed that Parabacteroides goldsteinii also plays a significant role in anti-obesity effects [88]. We found that PS supplementation caused a notable decrease in the abundance of genus Parabacteroides in mice fed a HFD, indicating that the supplementation of Parabacteroides spp. may be another effective approach to alleviate the adverse health effects induced by PS. Further research is necessary to gain a better understanding of the potential therapeutic applications of Bifidobacterium spp. and Parabacteroides spp. in managing PS-induced obesity and related metabolic disorders. Taken together, microbial dysbiosis is associated with PS-induced obesity.
HFD could significantly reduces the thickness of the intestinal mucus layer through microbial dysbiosis [77]. The mucus layer, which is formed by mucin proteins and hydrated gel with glycosylation, plays an essential role in separating the intestinal epithelium from its contents [89]. It prevents microorganisms from directly contacting epithelial cells and makes the intestinal mucosal barrier essential for human health. Dysfunction in this barrier has been linked to numerous diseases such as inflammatory bowel disease, microbial infections, and metabolic syndrome [90]. Therefore, we hypothesized that PS-induced microbial dysbiosis exacerbated mucosal layer injury. This study discovered that exposure to PS resulted in a reduction of mucus contents in the colon since it down-regulated MUC2 expression along with the tight junction-related genes such as ZO-1, Occludin, and Claudin-3 in HFD-fed mice. As a consequence, the destruction of the mucus layer selected and enriched certain bacteria like Lachnospiraceae, residing in the inter-fold regions of the colon, come in direct contact with the intestinal epithelium, thus initiating inflammation [91]. Previous study have shown that PS exposure induced systemic inflammation and thus induced insulin resistance [49]. Based on these results, we speculated that PS impaired the integrity of intestinal barrier to result in intestinal inflammation, which allows toxic molecules entry into the systemic circulation, thereby triggering systemic inflammation. This hypothesis was supported by the observation of reduced colon length, and up-regulated mRNA expression of inflammatory cytokines in the colon (TNF-α, IL-17A) and inguinal fat tissues (IL-17A), which was further bolstered by elevated serum levels of pro-inflammatory cytokines (TNF-α, IL-17A, IL-1β, and IL-6) and decreased serum levels of anti-inflammatory cytokines (IL-10). Furthermore, correlation analysis also revealed significant associations between the dysregulated gut microbiota induced by PS and indicators related to obesity, MUC2 expression levels, as well as inflammatory factors. Our findings suggest that PS-induced dysbiosis disrupts the gut microbiota, leading to increased penetration of toxic gut molecules into the body via impaired intestinal permeability in HFD-fed mice. This process drived a state of chronic low-grade inflammation that ultimately contributes to the development of obesity. These findings provide further evidence for the role of PS-induced microbial imbalance in perpetuating metabolic dysfunction, and highlight the potential utility of targeting the gut microbiota as a therapeutic strategy for treating obesity and associated disorders.
Alongside germ-free animal models, antibiotic-treated animal models are frequently employed in the investigation of intestinal bacteria [92]. To verify whether the adverse effect of PS on obesity is mediated by the intestinal flora, In this study, a combination of four broad-spectrum antibiotics - namely aminoglycoside, penicillin, neomycin, metronidazole and vancomycin were simultaneously administered for 8 weeks, resulting in a 97% reduction of commensal bacteria within murine intestinal microbiota. This investigation exhibitd certain limitations, most notably our inability to fully eradicated the intestinal microbial population. The findings indicated that antibiotic cocktail effectively resolved PS-induced obesity, thus supporting the hypothesis that gut microbiota play a crucial role in the development of this disorder. A more concerning result was that PS reduced the thickness of the mucus layer depended on the presence of gut flora. Therefore, our results indicated that PS harmed the intestinal barrier by affecting intestinal flora metabolites, leading to a low-grade inflammation that promotes HFD-induced obesity.
Moreover, we discovered that the PS induced low-grade inflammation and obesity were also dependent on intestinal flora. Notably, we found that the genus Desulfovibrio, a sulfate-reducing bacteria (SRB), exhibited an increased abundance in the PS + HFD group. This bacteria has been proven to produce LPS, which possesses 1000-fold endotoxin activity in comparison to other gut bacteria [48]. We also found that family Deferribacteres enriched at higher taxonomic levels such as phylum, class, and order in HFD + PS group. Mounting evidence suggests that family Deferribacteres were strongly associated with upregulated pro-inflammatory cytokines and exacerbation of inflammation and obesity-induced metabolic disease T2D [5, 93]. Furthermore, we observed a decrease in the number of family Bifidobacteriaceae, which is known to have a negative correlation with intestinal permeability, metabolic endotoxemia, and low-grade inflammation in the PS + HFD group. These shifted in gut microbiota induced by PS contributed significantly to the reduced expression levels of Muc2 and tight junction related genes, resulting in damage to the intestinal barrier function and induction of low-grade inflammation in mice colon tissue. We also found variations in the PS-induced microbiota alterations between the HFD-fed and ND-fed groups. However, we attribute this to the influence of the HFD on gut flora as it differs from ND. Nevertheless, ABX treatment did not completely eliminate lipid accumulation induced by an HFD compared to the ND diet, which indicated that gut microbes participated in HFD-induced obesity but were not the sole determining factor. Additionally, FMT exacerbated HFD-induced obesity, further supporting our conclusion that PS induced mucosal layer injury, low-grade inflammation and obesity depended on intestinal flora. Thus, microbiota dysbiosis and the damage of intestinal barrier collaborate to promote low-grade inflammation, which has been known to boost obesity development [94].
Obesity and related chronic inflammatory diseases have become a serious health problem both in developed and developing countries [95]. Recent studies indicate that the relationship between obesity, gut barrier, intestinal microbes, and their interactions is interlinked and chiefly dependent on dietary habits [96]. Our study demonstrated that frequently ingested PS exacerbated HFD-driven destruction of the mucus layer, inflammation, and obesity by altering the gut microbiome. We observed that PS amplified HFD-induced microbiota dysbiosis, demonstrating a superimposed effect between PS and HFD on intestinal integrity and flora. This highlights the importance of considering the impact of intestinal health on individuals with different dietary habits, particularly those consuming western diets, as PS may enhance the progression of obesity via alterations in the microflora.
{kind=link}