In the current study, we demonstrated that bilateral i.n. infusion with MPTP did not influence motor function of mice but impaired olfactory recognition and social memory consolidation. In addition, MPTP infusion induced compulsive- and anxiety-like behaviors. These NMS were associated with astromicroglial activation and changed levels of inflammatory mediators in the SNpc and striatum prior to the development of motor symptoms.
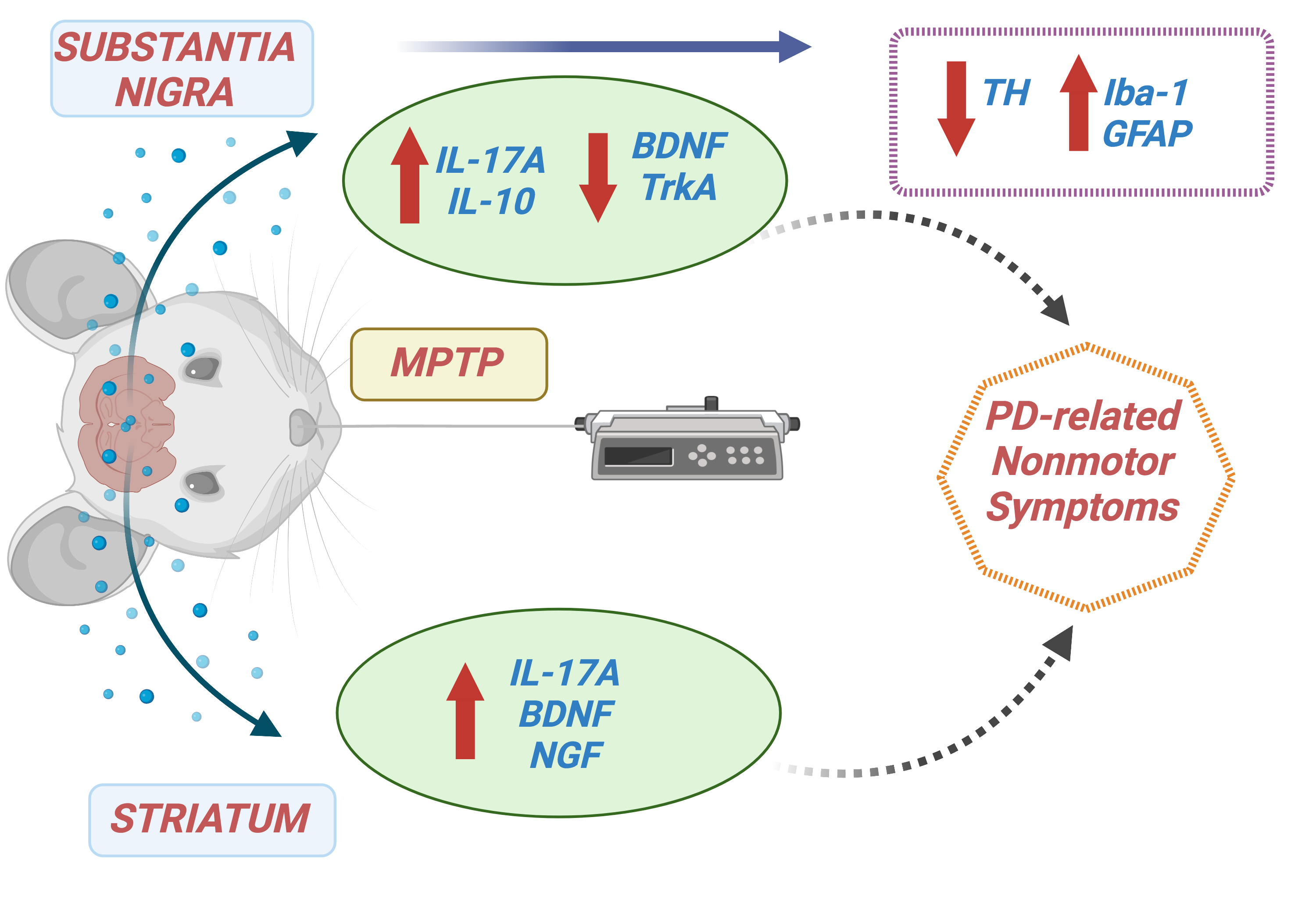
Loss of dopaminergic neurons in the SNpc can promote NSM. These neurons produce TH, an important enzyme in the dopamine production cascade, converting tyrosine to DOPA (Dunkley and Dickson 2019). We showed decreased neuronal immunostaining for TH in the SNpc of animals treated with a single low dose of MPTP with preserved motor function. Considering that the first motor symptoms of PD can appear decades after the onset of dopaminergic cell loss and NMS, our results suggest that the acute MPTP parkinsonism model can mimic the prodromal stage of PD (Schapira et al. 2017).
The diagnosis of PD relies on motor deficits, including bradykinesia, rigidity, and resting tremor. These motor features result from the loss of dopaminergic neurons in the SNpc (Berardelli et al. 2013). However, PD is associated with a series of remarkable NMS that can precede the motor symptoms. Herein, we demonstrated a decrease in social memory consolidation on the 6th day after MPTP i.n. infusion, using olfactory recognition test. Similar results were observed by Prediger and colleagues using the same murine model (DS Prediger et al. 2011). Hyposmia is present in more than 90% of patients with PD. The presence of hyposmia can be regarded as a biomarker for early premotor PD, particularly if it is combined with other early symptoms, such as cognitive dysfunction (Gaenslen et al. 2014; Doty 2012; Baba et al. 2011). Impaired olfaction following i.n. administration of MPTP could simply reflect damage to the olfactory epithelium (Kurtenbach et al. 2013). However, previous studies with MPTP found hyposmia without damage to the olfactory epithelium, i.e. the cause of MPTP-related olfactory impairment is still controversial (Dluzen 1992). In the open field test, MPTP treated mice spent less time in the center of the arena compared with controls, indicating anxiety-like behavior. Bilateral 6-OHDA-induced injury of the nigrostriatal pathway in rats promoted anxiety-like behavior associated with reductions in dopamine (Tadaiesky et al. 2008). In addition, individuals with anxiety have a higher risk of developing PD (Lin et al. 2015). Anxiety affects up to 60% of patients with PD and, often in association with depression, can occur before the onset of motor signs of PD.
The neuroanatomical and pathophysiological bases of nonmotor abnormalities in PD are complex, seem to involve non-dopaminergic pathways and remain largely undefined (Schapira et al. 2017). Studies using animal models have supported that neuroinflammation can increase dopaminergic neurodegeneration in PD. Microglial activation and increased levels of inflammatory mediators in the SNpc and striatum have been related to MPTP administration in mice (Tristao et al. 2014; Machado et al. 2016; Mustapha and Taib 2021). Interestingly, Yasuda and colleagues reported that MPTP-sensitive C57BL/6 mice showed increased inflammatory cytokine levels in the cerebrospinal fluid, while it was not observed in MPTP-resilient BALB mice (Yasuda et al. 2008). These results indicate that the neuroinflammatory response might trigger the susceptibility of C57BL/6 mice to MPTP-induced loss of dopaminergic neurons. Studies of inflammatory response in the early stage of parkinsonism syndrome (i.e., before the motor symptoms occurrence) are still missing (Yasuda et al. 2008). In the current study, we demonstrated increased immunostaining for Iba-1 and GFAP in the SNpc of MPTP-treated mice compared to control mice previously to the motor symptoms, corroborating the hypothesis of an early role of astromicrogliosis in PD pathophysiology (Tristao et al. 2014).
MPTP intoxication induces neuronal loss, triggering microglial activation and proliferation and releasing inflammatory mediators that contribute to neurodegeneration in a cyclical process (Machado et al. 2016). Alongside microglial activation, we found MPTP-induced increase of nigrostriatal levels of IL-17A. Similarly, increased brain levels of IL-17A following blood brain-barrier disruption were observed in MPTP-treated mice. In an in vitro study, IL-17A induced microglial activation, death of TH-positive neurons, and decreased dopamine levels. Interestingly, IL-17A exacerbated dopaminergic neuronal loss only in the presence of microglia, and silencing the IL-17A receptor gene in microglia abolished the IL-17A effect (Liu et al. 2019). In line with this, Ginkgolide K had a dopaminergic neuroprotective effect in MPTP-treated mice, decreasing microglia-mediated inflammation and consequent infiltration of peripheral CD4+IL-17+ T cells into the mice’s brain (Miao et al. 2022). These founds suggest that, by activating microglia, IL-17A indirectly potentiates dopaminergic neuronal death, since microglia definitely express IL-17R and play a pivotal role in neuroinflammation and neurodegeneration in PD (Garcia-Esparcia et al. 2014; Liu et al. 2019). Astrocytes also respond to IL-17 through generating chemokines to promote the recruitment of peripheral leukocytes. In addition, IL-17 reduces the ability of astrocytes to metabolize glutamate, inducing glutamatergic excitotoxicity (Chen et al. 2020). We also observed a higher concentration of IL-10, an anti-inflammatory cytokine, in the MPTP group compared to the control. The transfection of Treg cells and the consequent increase of IL-10 levels into the brain of MPTP-treated mice decreased neuronal loss and microglial activation, as well as increased production of both BDNF and GDNF, suggesting a neuroprotective role for IL-10 in the MPTP model (Reynolds et al. 2007). Therefore, the elevation of IL-10 in the SNpc might be an attempt to counterbalance the proinflammatory effects of increased astromicrogliosis and IL-17 levels (Kwilasz et al. 2015).
In the current study, we also demonstrated the reduction of BDNF levels and mRNA expression of TrkA in the SNpc of MPTP-treated mice. BDNF is involved in the activity and survival of dopaminergic and motor neurons. Cell-mediated delivery of BDNF increased dopamine levels, and nigral infusion of BDNF reversed the reduction of dopamine in the murine MPTP model (Hung and Lee 1996; Isacson et al. 1995). In addition, BDNF gene ablation led to impaired striatal development and severe motor dysfunction in mice (Li et al. 2012). BDNF and NGF concentrations were also decreased in the nigrostriatal system and serum of patients with PD (Chmielarz and Saarma 2020; Nasrolahi et al. 2018; Scalzo et al. 2010a). Although NGF levels remained similar to those in the control group, the MPTP-induced reduction of mRNA TrkA expression may inhibit the neuroprotective effects of this neurotrophic factor. TrkA is the receptor most related to NGF and its regulation in the CNS (Nasrolahi et al. 2018). Conversely, in the striatum of MPTP-treated animals, we demonstrated increased levels of NGF and BDNF. Once again, this may be an attempt to counterbalance the neuroinflammatory and neurotoxic response induced by MPTP intoxication in the early-stage model of PD.
In conclusion, our results suggest that MPTP can induce neuronal loss and neuroinflammation causing a series of cognitive and behavioral changes in the early stage of experimental PD. Our data also indicate that acute MPTP intoxication by i.n. route represents a valid model of prodromal PD.
{kind=link}