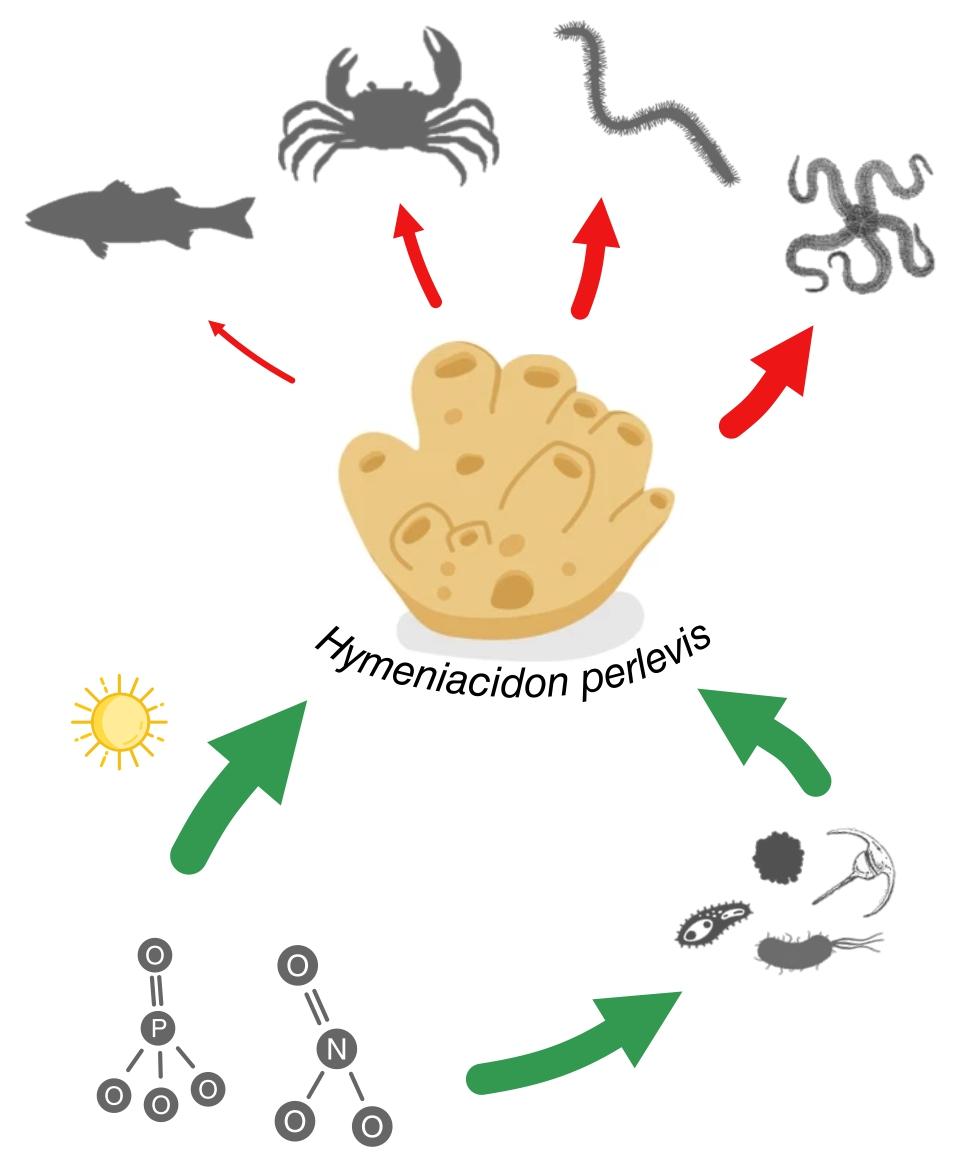
In this study, we found that the individual growth and abundance of H. perlevis was relatively higher in the SAO channel -where anthropic nutrient loads are higher- compared to the CTL channel. The bottom-up variables that better explained the abundance pattern of the sponge were the concentration of nitrate, phosphate and total suspended solids. We also found that some crabs, fishes, polychaetes and ophiuroids would be potential consumers of H. perlevis. However, the effects of consumers were negligible on the individual growth dynamics of the sponge, even though some benthic invertebrates (polychaetes and ophiuroids) of the CTL channel would be exerting the greatest consumption pressure over the sponge. These findings support our main hypothesis that the population of H. perlevis in San Antonio Bay is mainly bottom-up controlled, based on the concentration of trophic resources.
Previous studies addressing the "top-down and bottom-up" dichotomy in sponges have been conducted exclusively in tropical coral reefs, where changes in sponges density or biomass may affect the development and maintenance of corals (see Wulff 2012, 2017; Pawlik et al. 2018). Furthermore, in temperate environments the variability of their physical (e.g, solar radiation, temperature, duration and intensity of stratification of the water column) and biological conditions (e.g, changes in primary productivity) over wide spatial and temporal scales, hinders the generalization about the most influential factors in the regulation of populations (Schiel 2004; Lesser 2006). Through the prolific literature on the top-down and bottom-up regulation and their interaction in filter feeders on temperate coasts, (see Menge 1992, 2000; Menge et al. 1997), this is the first study to assess the relative contribution of top-down and bottom-up controls on the regulation of a sponge population, in a temperate environment.
Here, we found that changes on the individual growth of H. perlevis transplants during the experiment are consistent with seasonal growth dynamics observed at temperate environments (Cao et al. 2007; Gastaldi et al. 2020). However, while transplants increased in surface area and volume, they decreased in dry weight. Such inconsistencies between variables may be related to specific growth mechanisms that sponges develop after a disturbance on its somatic structure (Ayling 1983) and have already been observed in H. perlevis in the SAO channel (see Gastaldi et al. 2017). In general, during the first two–four weeks after tissue cuts, sponges show morphological and structural changes involving instances of cellular disorganization and transformation, differentiation and cell rearrangement, where transplants may lose weight (de Caralt et al. 2008) although this is not necessarily reflected in a decrease in its volume or surface area given the three-dimensional rearrangement of its structure (Ayling 1983).
During the experimentation, we found that channels with contrasting trophic resource concentration affect the growth of H. perlevis only during winter-spring. This suggests that the availability of trophic resources has a differential effect on individual sponge growth, probably related to its physiological state and annual growth cycle. Channels with different concentrations of resources had significant effects during the season with lower growth rates of the sponge, since after winter dormancy, the sponge begins to grow in spring (Cao et al. 2007; Gastaldi et al. 2020). In this sense, a greater subsidy of trophic resources in the SAO channel at the beginning of the growth season could favorably stimulate its growth. On the other hand, this does not happen during the autumn, when the sponge grows more than in winter-spring. Although autumn is not the season when H. perlevis reaches its greatest individual growth (see Cao et al. 2007; Gastaldi et al. 2016, 2020), it seems that trophic resources are not the determining factor for the sponge growth. It is likely then, that other variables than trophic resources would best explain sponge growth dynamics, while low levels of resources are sufficient for its growth and maintenance. During the autumn, other abiotic factors could become more relevant. For instance, in SAB, the population abundance of the sponge seems to be strongly related with seawater temperature (Gastaldi et al. 2016, 2017).
Regarding consumers, we found some taxa might be potential consumers of H. perlevis. However, consumers were not relevant controlling the individual growth of the sponge, in any of the experimental instances. Therefore, temporal variations in the abundance of predators -if any- or in their consumption rate would potentially result in a negligible effect on the sponge growth. It is also possible that the mesh used to exclude consumers in the consumer exclusion cage was not effective enough to avoid small organisms. However, we did not observe bite marks on the surface of the sponge nor the presence of organisms inside the consumer exclusion cages during the weekly cleaning of the experimental units. We found that except for the occurrence of the unidentified polychaete in both channels, the rest of the potential consumers of H. perlevis when they were collected in one of the channels did not appear in the other, and vice versa (see Table S1). This could be due to the fact that the communities in both channels differ, maybe related to the anthropogenic nutrients input (see Martinetto et al. 2010). The methodology we used to identify the potential consumers of H. perlevis is limited. The mere presence of spicules inside an organism is not absolute evidence of their consumption, because spicules are often found in sediment and debris (Guida 1976). However, the setting of the conservative threshold of 50% of incidence strengthens the identification criterion. Furthermore, we found that only in some of the potential consumers identified here there were also spicules of other sponge species from the bay, although in lower abundances than H. perlevis (Gastaldi 2016). These findings allow us to assume that the identified potential consumers are eating sponges. According to this, we found spicules of the sponges Pachychalina tenera, Halichondria elenae, and Cliona celata in the fish Odontesthes platensis and the crabs Cyrtograpsus affinis, C. angulatus and Neohelice granulata.
Contrasting conclusions have been reported from top-down and bottom-up studies on tropical coral reef sponges. Some studies conclude that there is no evidence of a bottom-up control, and that individual sponge growth is only top-down controlled (Pawlik et al. 2013, 2015). These studies highlight that in tropical coral reefs there is no food limitation for sponges (either POC or DOC) that could lead to a bottom-up control (Pawlik et al. 2015, 2018). On the other hand, other studies agree on the relevance of bottom-up control on sponges (Lesser 2006; Trussell et al. 2006; Lesser and Slattery 2013, 2018; Wulff 2017). For instance, a recent study showed that several sponge species from coral reefs -when consumers are excluded- reach higher growth when transplanted to sites with higher concentrations of picoplankton and dissolved nutrients, while their growth is not affected by predation when they are incubated in their original site (Wulff 2017). Our study, the first carried out in a temperate system, reveals that H. perlevis growth is mostly bottom-up controlled. At the end of the experimental study, transplants were bigger when incubated in the channel with higher concentration of trophic resources, at least during the winter-spring. Despite the presence in both channels of several consumers of H. perlevis, these did not have a significant effect on the growth of the sponge. Therefore, in SAB, the top-down control over H. perlevis growth would be negligible compared to the bottom-up control.
A greater abundance of H. perlevis was also recorded in the channel with the highest concentration of trophic resources and, among the bottom-up variables considered, those best explaining the variability of the sponge cover were the concentration of nitrate, and to a lesser extent, phosphate and total suspended solids. Some authors have already found a positive correlation between the abundance of sponges and the concentration of trophic resources (Bell 2008b; Wulff 2012). These relationships are associated with the concentration of both DOC and POC (Reiswig 1971; see Lesser and Slattery 2013; Wulff 2017; Pawlik et al. 2018; Pawlik and McMurray 2020). DOC accounts for a large part of the sponge diet (Pawlik et al. 2018; Pawlik and McMurray 2020), for example, ∼ 70% of the diet of the giant barrel sponge Xestospongia muta is based on DOC (McMurray et al. 2016). Unfortunately, here we neither measured DOC concentration nor its relationship with the abundance of H. perlevis in SAB. Future studies should consider the analysis of DOC concentration and its contribution to the growth and abundance of the sponge. On the other hand, sponges can actively filter up to 75–99% of particulate suspended solids (~ POC, Ribes et al. 1999). For example, the abundance of Cliona delitrix on coral reefs increases with the concentration of suspended organic matter along a gradient of a sewage discharge (Chaves-Fonnegra et al. 2007). Likewise, at the Caribbean coral reefs, an increase in density of Callyspongia vaginalis along a depth gradient is driven by a higher concentration of picoplankton in deeper waters, mostly phytoplankton, prochlorophytes and heterotrophic bacteria (Lesser and Slattery 2013). According to this, in this study we found that in SAB, the variability in the cover of H. perlevis could be related to the availability of total suspended solids as a proxy of POC concentration.
A positive correlation between the abundance or biomass of sponges and the concentration of nutrients (dissolved inorganic nitrogen and phosphorus) at enriched environments is mainly attributed to the rapid intake of nutrients by bacterioplankton or phytoplankton, leading to a greater availability of particulate trophic resources for sponges (Reiswig 1971, 1974; see Holmes 2005; see Ramsby et al. 2020) or POC. In SAB, the concentrations of nutrients were significantly higher in the SAO channel than in the CTL channel (by two orders of magnitude higher for nitrate). Here, the difference in the concentration of nutrients is due to an anthropogenic supply through both point sources (fish processing factories) and diffuse sources (septic system) of sewage (Teichberg et al. 2010; Martinetto et al. 2011; Fricke et al. 2016). Since sponges harbor photosymbionts that fix nutrients into complex carbon compounds (photosynthates) that are then transferred to the sponge (see Cheshire and Wilkinson 1991; Weisz et al. 2010; see Freeman and Thacker 2011), a higher nutrient concentration would affect the abundance and growth of sponges via the autotrophic pathway (Reiswig 1981; de Goeij et al. 2008; Wulff 2012). The variation in the abundance pattern of H. perlevis in relation to the concentration of nitrate, phosphate and total suspended solids suggests that the sponge could be supporting its nutritional requirements, not only by a heterotrophic pathway but also by an autotrophic pathway, through its photosymbionts. For example, Aplysina cauliformis, a Caribbean sponge, obtains up to 75% of its energy reserves from photosymbionts (Freeman and Thacker 2011). However, the role that H. perlevis symbionts (e.g., cyanobacteria; see Alex et al. 2012) may have on the nutritional requirements of the specie is still unknown.
In summary, a higher concentration of trophic resources was recorded in the channel next to the city. There, both the abundance and individual growth of H. perlevis were greater, increasing its cover along with the concentration of trophic resources. Contrary to our prediction, the greatest consumption pressure was exerted by the potential consumers of the channel isolated from the city, where the concentration of trophic resources is lower, while in both channels, the effect of predation -if any- on the individual sponge growth was masked by the bottom-up control.
{kind=link}