The level and duration of ATOH1 transgene expression determine supporting-cell-to-hair-cell conversion in the adult mouse utricle ex vivo
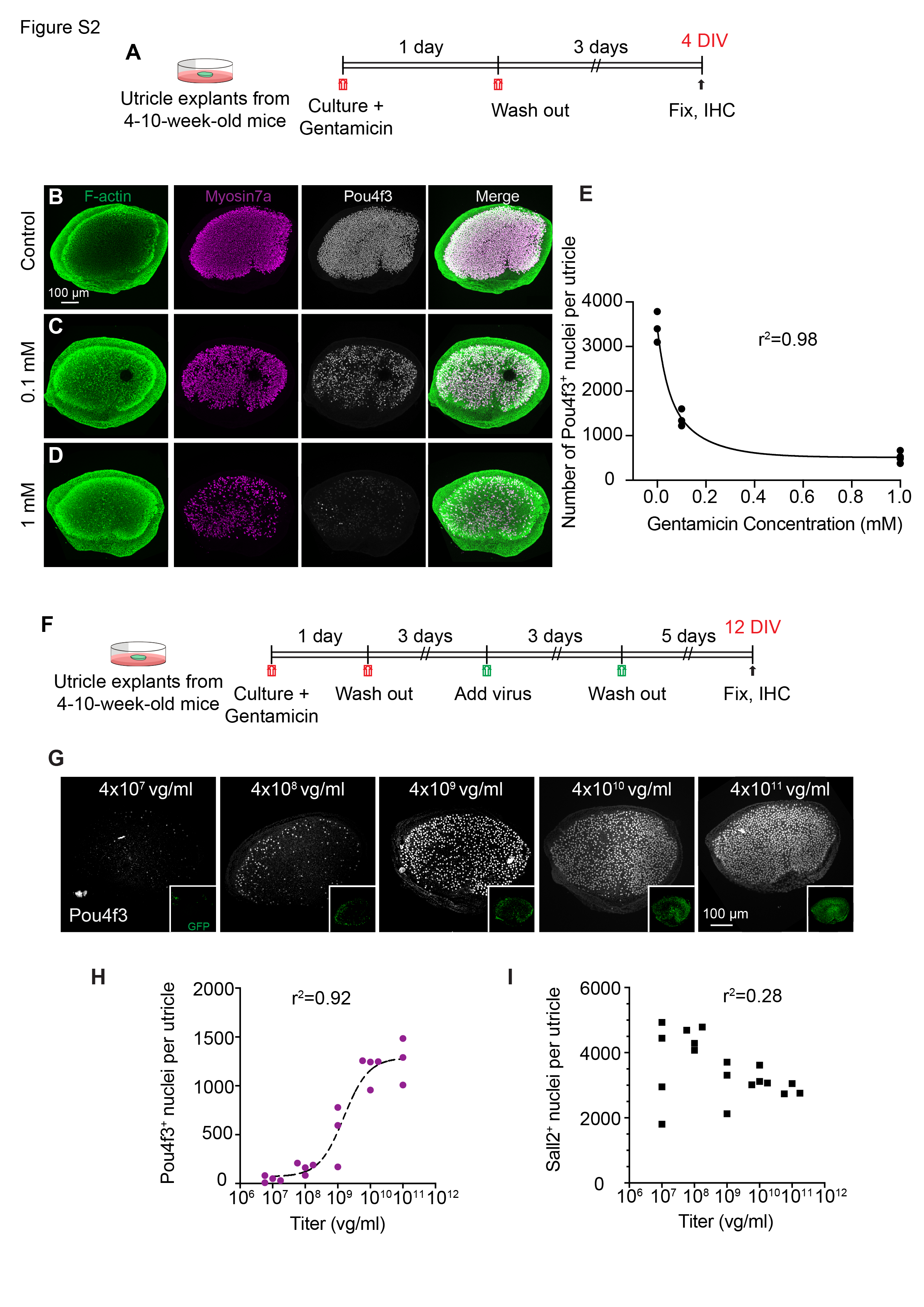
Adenoviral-based overexpression of mouse or human ATOH1 converts supporting cells into new hair cells in cultured utricles from adult mice, rats, and humans14,21,33,34. Early work comparing promoters delivered via adenoviral vectors concluded that lower levels of ATOH1 expression resulted in more regenerated hair cells compared to higher levels34. To begin to determine how spatiotemporal expression of ATOH1 affects hair cell regeneration, we cultured adult mouse utricle, lesioned hair cells with the aminoglycoside gentamicin, and subsequently transduced them with escalating titers of AAV-CMV-ATOH1 (Fig. S1, S2A-F). We observed a strong, dose-dependent conversion of supporting cells to Pou4f3 + hair cells (Fig. S2G-H), consistent with previous results 20.
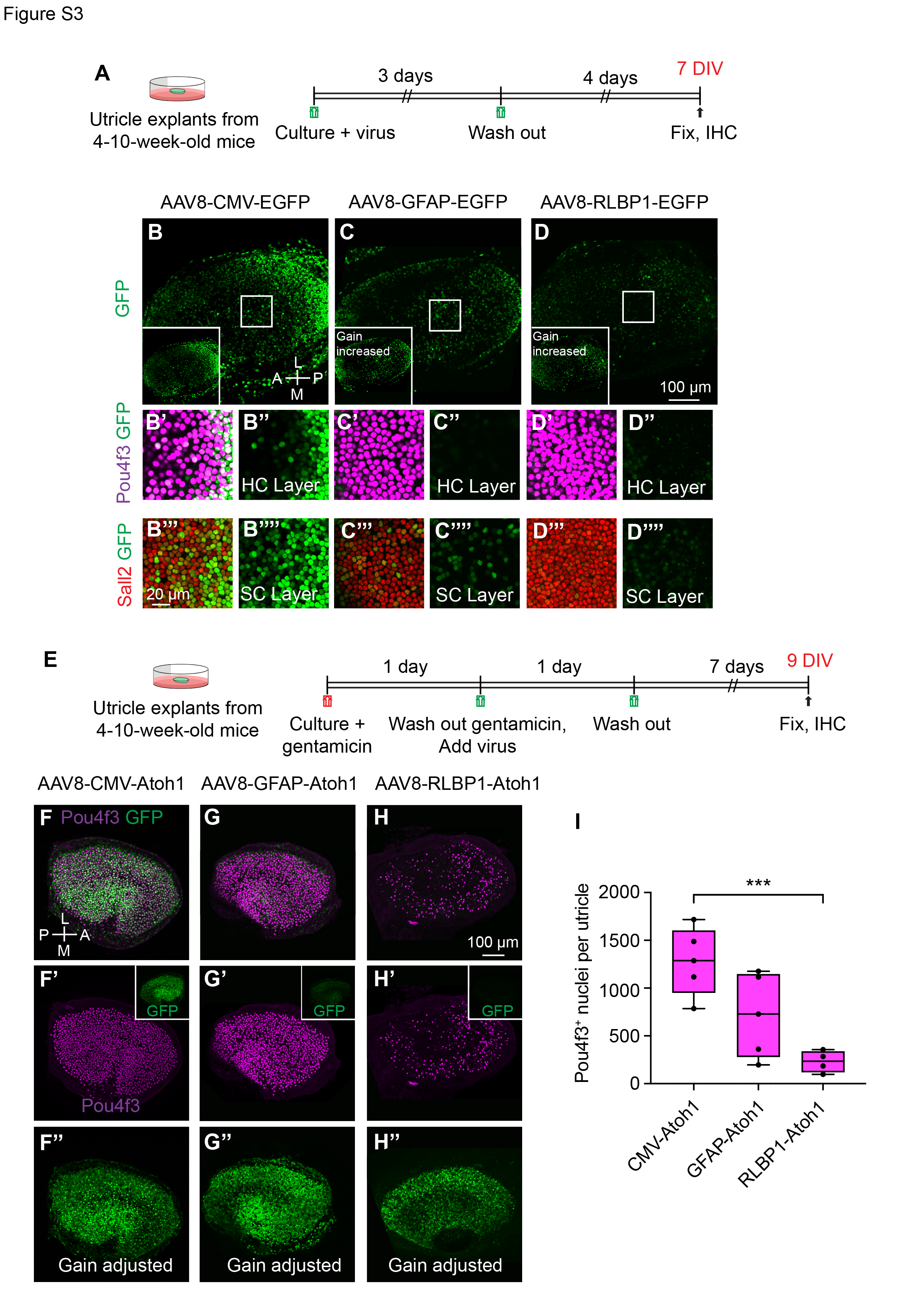
Higher titers of AAV increased the total number of supporting cells transduced (Fig. S2G-I), and may contribute to the efficiency of AAV-CMV-ATOH1 in inducing hair cell conversion. We reasoned that gene regulatory elements (e.g., promoters) that exhibit differing strengths and are specific to supporting cells may allow titration of the level of ATOH1 transgene expression at equivalent AAV doses. To identify such regulatory elements, candidate sequences were identified from single-cell genomics data 35, cloned into AAV constructs driving an H2B-EGFP transgene (Fig. S1), and tested in adult utricle explants at similar doses (AAV8-CMV-H2B-EGFP (8.8x1011 vg/ml), AAV8-GFAP-H2B-EGFP (1.52x1013 vg/ml), or AAV8-RLBP1 -H2B-EGFP (8.6x1012 vg/mL)). We selected the AAV8 serotype as the vector because it has been shown to transduce utricular hair cells and supporting cells at high rates in vivo20,36. We transduced utricle explants from adult mice with AAV8 carrying the regulatory sequences associated with cytomegalovirus (CMV), glial fibrillary acidic protein (GFAP), and retinaldehyde binding protein 1 (RLBP1) for 4 days and found decreasing levels of EGFP intensity within supporting cells among these promoters (Fig. S3A-D’’’’, S5A). When ATOH1-GFP under different promoters was delivered, the CMV promoter induced relatively higher ATOH1-GFP intensity than GFAP and RLBP1 (1.5 and 6.6-fold), and GFAP was 4.4-fold more intense than RLBP1 at 12 days ex vivo. These differences increased to 2.2, 12.1, and 4.4-fold, respectively, at 20 days ex vivo (Fig. 1I). Furthermore, the CMV promoter drove robust EGFP expression in both hair cells and supporting cells, while with GFAP and RLBP1 expression was mostly restricted to supporting cells, suggesting that these regulatory elements are not active in hair cells (Fig. S3B-D’’’’).
Having identified a set of regulatory elements driving varying degrees of EGFP expression, we next tested the relationship between levels of ATOH1 transgene expression and hair cell regeneration. Gentamicin-damaged utricle explants37,38 were transduced with AAV8-CMV-ATOH1-H2B-EGFP (5.7x1012 vg/ml), AAV8-GFAP-ATOH1-H2B-EGFP (7.2x1012 vg/ml), or AAV8-RLBP1-ATOH1-H2B-EGFP (1.4x1013 vg/mL) and then cultured for an additional 4–13 days to allow time for hair cell regeneration to occur (Fig. 1A-G, S3E-H’’). Regeneration of hair cells was observed in all three treatment groups, and the level of regeneration increased in the order (CMV > GFAP > RLBP1) similar to their corresponding EGFP intensity (Fig. 1H-J, S3E-I). The number of hair cells per utricle at the end of the culture period were 1278.8 ± 355.1 for AAV8-CMV-ATOH1, 715.4 ± 437.7 for AAV8-GFAP-ATOH1, and 230.5 ± 113.3 for AAV8-RLBP1-ATOH1 (p < 0.05, one-way ANOVA; Fig. S2E, 3F-I.
To compare the extent of regeneration over time with each regulatory element, we separately assessed the number of hair cells in utricle explants at five and 13 days after AAV transduction. The mean number of hair cells per utricle increased significantly between the two timepoints for CMV- and GFAP-ATOH1, but not RLBP1-ATOH1 treatment (Fig. 1B-G, J), indicating that new hair cell generation was still occurring between the 2 timepoints. Furthermore, the number of hair cells remained higher for CMV than GFAP and RLBP1 at both timepoints (Fig. 1H-J). Taken together, these data suggest that CMV is a stronger promoter than both GFAP and RLBP1 and induced more hair cell regeneration in damaged mouse utricles ex vivo.
scRNA-seq reveals transcriptome-wide changes during AAV-ATOH1-mediated supporting cell-to-hair cell conversion
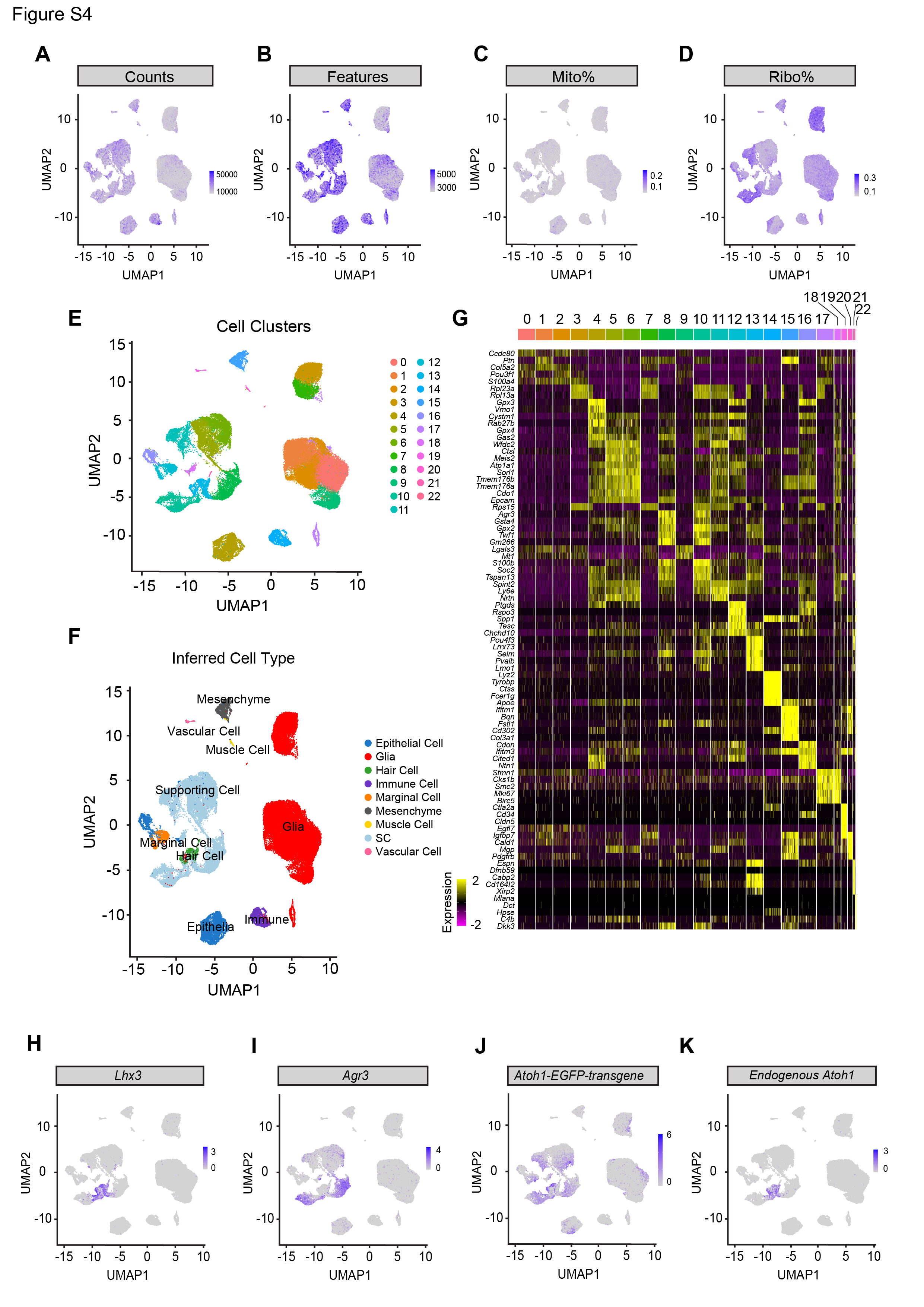
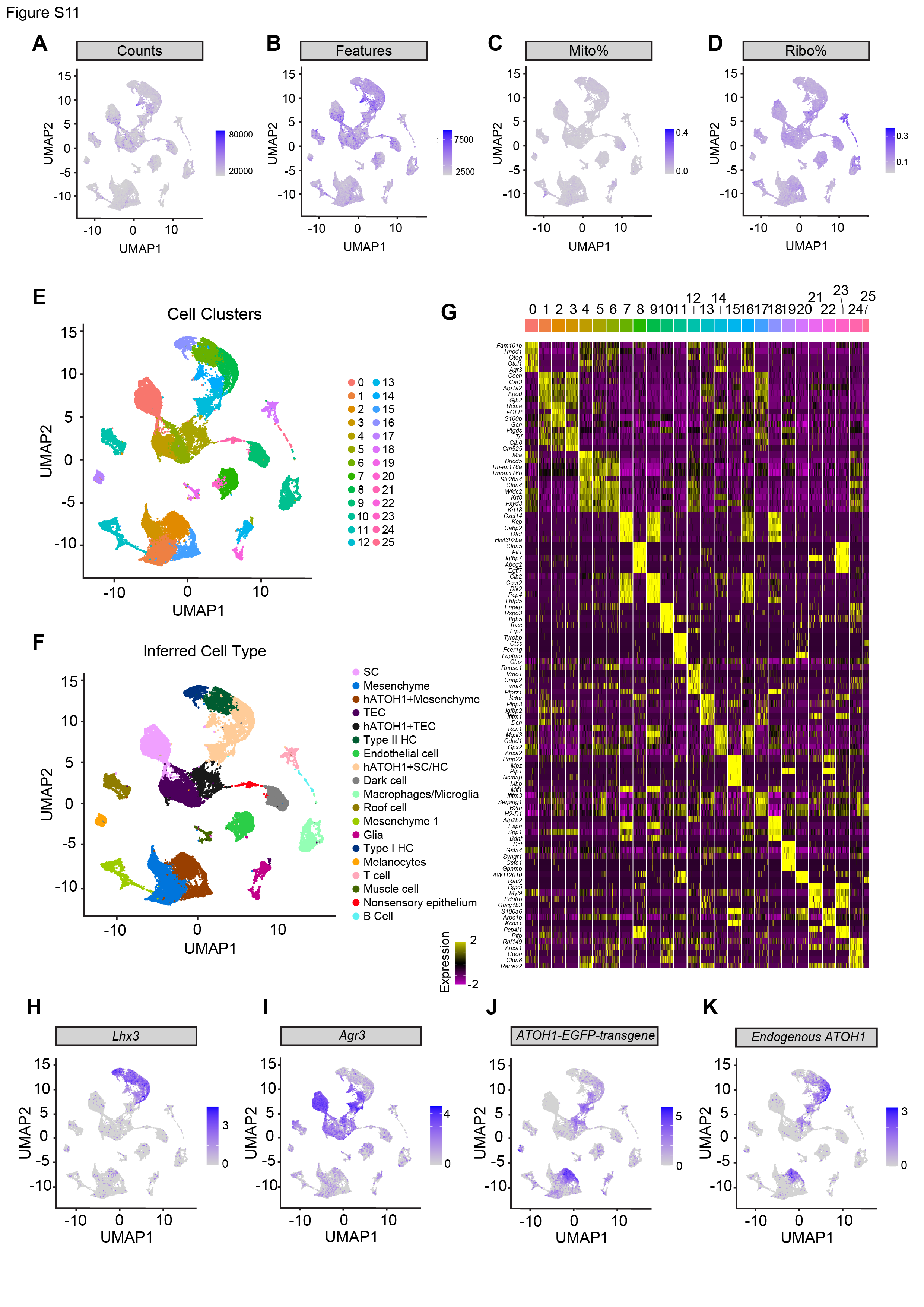
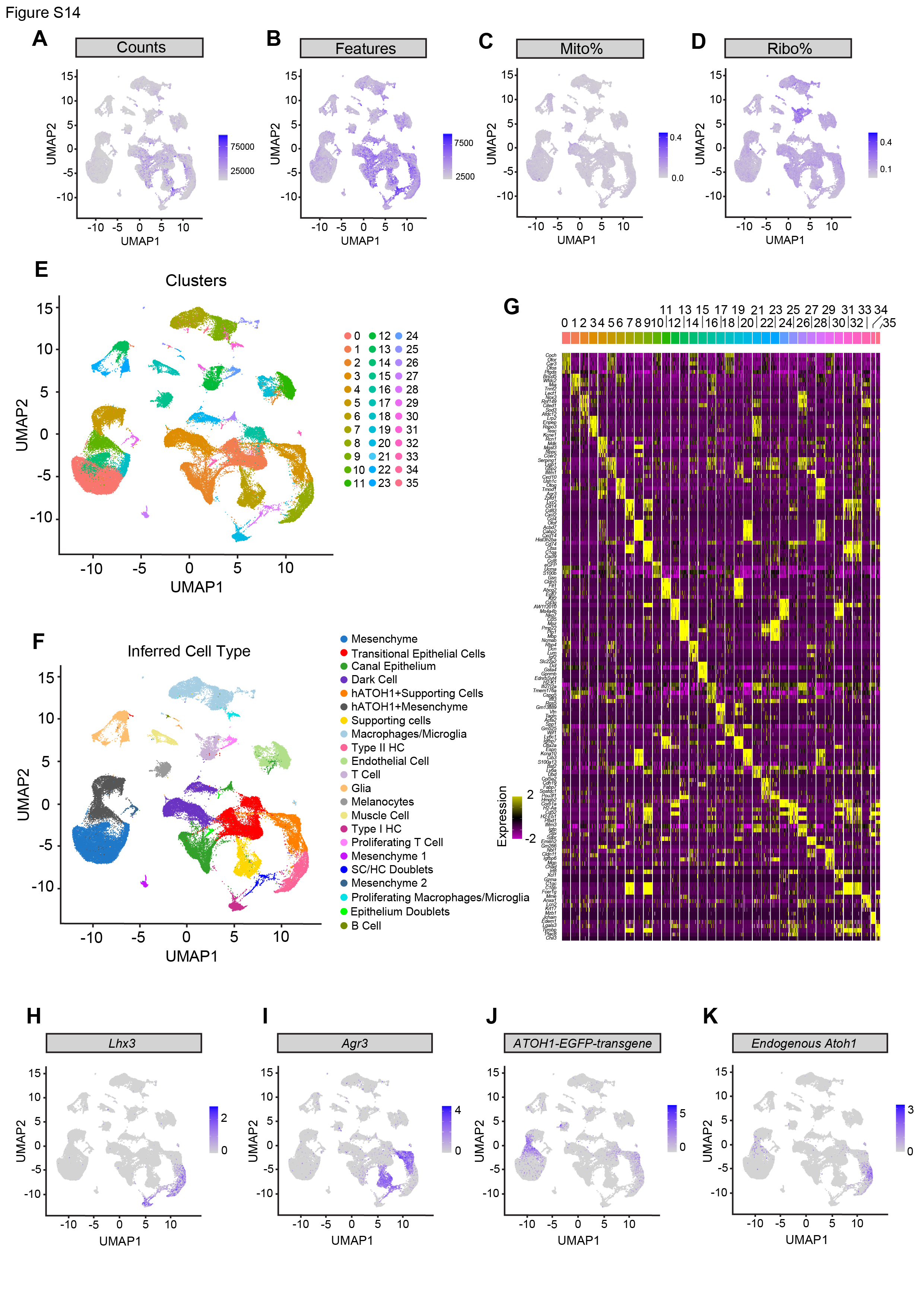
To map the transcriptomic changes during supporting cell-to-hair cell conversion in utricles treated with the ATOH1 transgene, gentamicin-damaged utricle explants were transduced with AAV8-CMV-ATOH1, AAV8-GFAP-ATOH1, or AAV8-RLBP1-ATOH1 and then were maintained in culture for 7, 12, or 20 days before being processed for single-cell RNA-Seq (scRNA-Seq; Fig. 2A). A total of 34,574, 30,428, and 21,891 high-quality cells were collected for each regulatory element, respectively (Fig. S4A-D). After merging these datasets and using marker gene expression to enrich supporting cells and regenerating hair cells (Fig. S4A-I), the final dataset consisted of 6,901 cells. UMAP and heatmap visualization of the merged dataset from cells taken at all timepoints revealed six distinct cell clusters that were separated based on gene expression into distinct phases of supporting cell-to-hair cell conversion (Fig. 2B-B’, D). The six clusters organized by differentiation status along an unbiased pseudotime gradient (Fig. 2E), with maturation of regenerated hair cells progressing over time in culture (Fig. 2G). We also found 1 clusters of immature and 2 clusters of mature regenerated hair cells, which we designed as hair cell a and β, suggesting a divergent of cell fates. Compared to 7 days in vitro (DIV) when only immature hair cells were present, more mature hair cells were observed at 12 DIV and 20 DIV (61.3% and 70.2% of HCs, respectively; Fig. 2G). While these six clusters were observed after transduction with all three regulatory elements, higher proportions of converting supporting cells and mature hair cells were found after treatment with ATOH1 driven by CMV and GFAP than by RLBP1 (Fig. 2F).
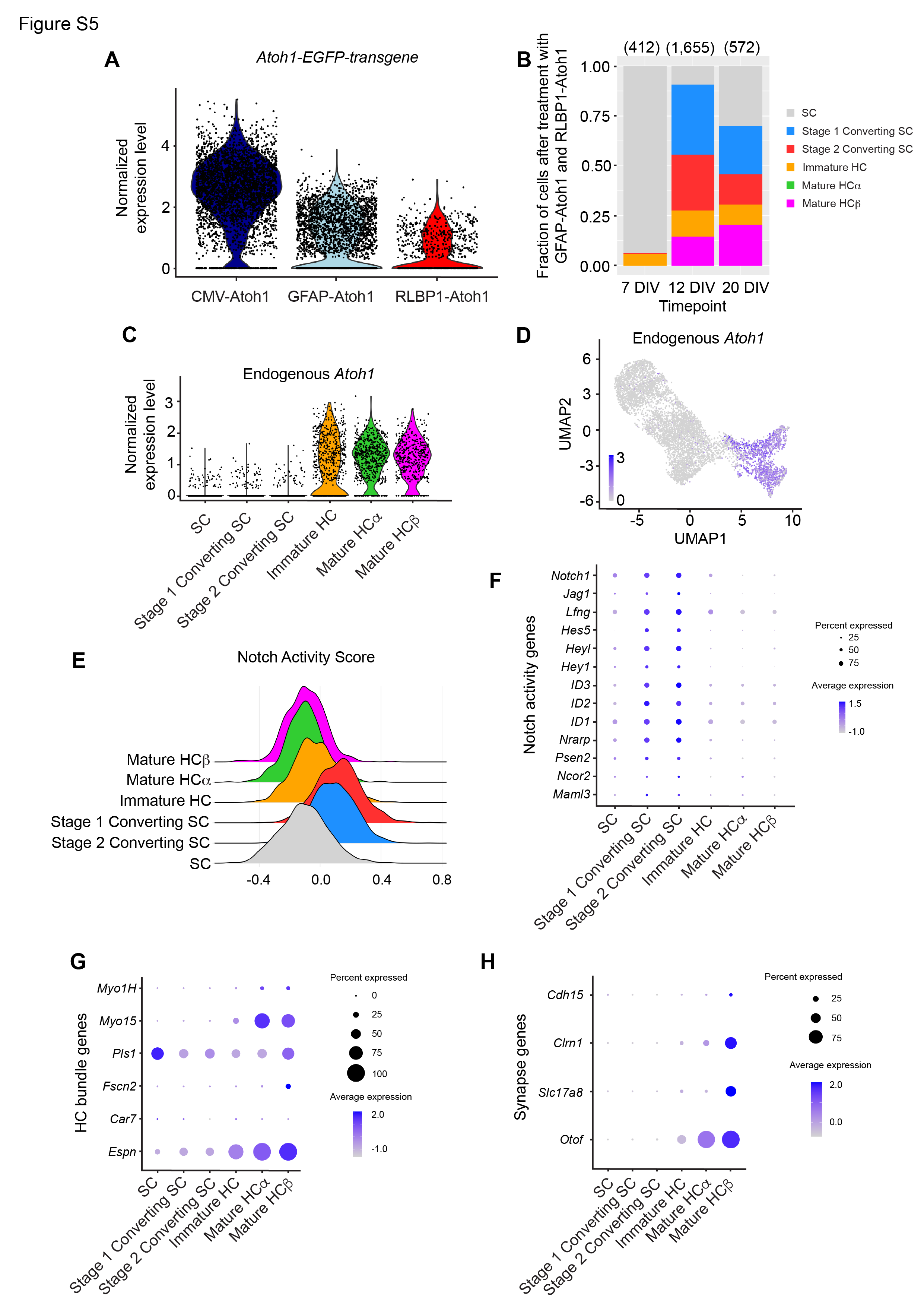
Supporting cells that responded to ATOH1 overexpression (stage 1 converting SC and stage 2 converting SC) initially downregulated known supporting cell genes (Otol1, Otog, Tspan8, and Gsn), and upregulated Notch pathway genes (Hey1, Hes5, Jag1, and Notch1) prior to the onset of expression of early hair cell genes (Pou4f3, Lhx3, Hes6, and Dlk2) (Fig. 3J, S5E-F and Table S1). A “Notch Activity Score” based on expression of 57 genes associated with the canonical Notch pathway (see Methods, Table S3) showed that Notch pathway activity is the highest in the stage 1 and 2 converting supporting cell clusters (Fig. S5E-F). As Notch signaling suppresses supporting cells from acquiring a hair cell fate39–41, these findings suggest that Notch gene expression becomes upregulated in converting supporting cells possibly as a result of exogenous Atoh1. At 20 DIV, there were substantially higher percentages of stage 1 and 2 converting supporting cells (Fig. 2G, S5B). Interestingly, many supporting cells did not show any signs of conversion despite displaying levels of ATOH1 transgene expression comparable to converting SCs, with 31.5% of SCs at 20 DIV remaining in the supporting cell cluster (Fig. 2G, 3A, Table S1). These ATOH1-transgene+, non-converting SCs exhibited lower Notch activity scores than stage 1 and 2 converting supporting cells (Fig. S5E-F).
Consistent with EGFP intensity corresponding to each regulatory element (Fig. 1B-F), the levels of ATOH1 transgene in supporting cells was similarly ordered (average across all cells, CMV = 18.6 normalized read counts; GFAP = 3.3; RLBP1 = 1.2; Fig. S5A). Moreover, the percentage of regenerated hair cells (immature hair cells, mature hair cell a and b) increased with regulatory elements associated with higher ATOH1 transgene expression level (CMV = 28.7%; GFAP = 24.9%; RLBP1 = 12.2%; Fig. 2F).
ATOH1 transgene expression under control of a supporting-cell-specific regulatory element becomes silenced in regenerated hair cells
Two of the regulatory elements used to drive ATOH1 expression, GFAP and RLBP1, led to limited or no EGFP expression in hair cells (Fig. 1B-F, S3F-H’’). Based on these results, ATOH1 transgene expression was expected to become downregulated as supporting cells convert into hair cells (Fig. 3A-D). ATOH1 transgene expression was detected in several clusters (supporting cells, stage 1 and 2 converting supporting cells, immature hair cells, and mature hair cells α) for all three regulatory elements utilized, indicating that they are active during the early stages of supporting cell conversion and hair cell specification (Fig. 3A). In contrast, the mature hair cell β cluster segregated into distinct groups visible on the UMAP based on regulatory element and ATOH1 transgene level (Fig. 2B-C, 3B). Hair cells from the CMV group almost exclusively comprised the mature hair cell α cluster (99.8% of cells), whereas the GFAP and RLBP1 groups comprised the mature hair cell β cluster (92.9% of cells; Fig. 2B-C). As predicted, the ATOH1 transgene was expressed at high levels in the mature hair cell α cluster, but it was significantly reduced or absent in mature hair cell β (Fig. 3A-B), suggested that it is downregulated as regenerated hair cell mature.
ATOH1 autoregulates its own gene expression by binding to regulatory regions in the 3’ enhancer of the endogenous gene42,43. We distinguished Atoh1 transcripts originating from the endogenous locus from vector transcripts via the WPRE element at the 3’ end of the ATOH1 transgene. Interestingly, expression of endogenous Atoh1 was detected in the immature hair cell, mature hair cell α, and mature hair cell β clusters, but not supporting cell or either converting supporting cell clusters (Fig. S5C-D). This finding suggests that ATOH1 protein produced from the vector transgene induced expression of endogenous Atoh1, and endogenous Atoh1 expression persisted in the mature hair cell β cluster after Atoh1 transgene expression from the GFAP and RLBP1 regulatory elements became silenced.
Silencing of AAV-Atoh1 transgene from supporting cell-specific regulatory elements promotes maturation of regenerated vestibular hair cells ex vivo
Atoh1 is expressed transiently in hair cells during development, and misexpression of Atoh1 in developing hair cells delays maturation44,45. We compared the transcriptomes of the mature hair cell α and β clusters to assess whether silencing of Atoh1 transgene expression driven by supporting cell-specific regulatory elements promotes maturation. As a measure of maturation, “hair cell maturity indices” were created based on the top 200 mature hair cell and 100 immature hair cell genes that are up- or down-regulated as hair cell differentiate during development (see Methods, Table S3) and were used to score cell clusters along the supporting cell-hair cell axis. The mature hair cell β cluster scored higher on the “Mature hair cell” and lower on the “Immature hair cell” indexes compared to mature hair cell α cluster (Fig. 3E-F), confirming that the former is more mature at the transcriptome level. The expression of select index genes is shown in Fig. 3I-J, highlighting the differentially expressed genes in the mature hair cell β cluster including those critical for synaptogenesis (Otof, Slc17a8), stereocilia formation (Espn, Fscn2), and mechanotransduction (Clrn1) (Fig. S5G-H).
Mammalian vestibular organs contain two types of hair cells, type I and type II, that are found throughout the organ, along with a central striolar region and a peripheral extrastriolar zone46,47. The functional significance of hair cell subtypes and zonal distribution have yet to be fully elucidated, but type I hair cells in the striolar region are associated with fast, phasic responses and type II hair cells with more static, tonic responses 48,49. To assess the extent regenerated hair cells differentiate into specialized hair cell subtypes, we developed maturation indices for type I and II hair cells based on the expression of genes which are upregulated in each subtype in the adult mouse utricle (see Methods, Table S3). We found that the mature hair cell β cluster scored higher on the type II hair cell index compared to the mature hair cell α cluster (Fig. 3G-H). Cells from both the mature HCα and mature HCβ clusters scored relatively low on type I hair cell index, indicating regenerated HCs were not adopting a type I fate ex vivo (Fig. 3H). These data indicate that the expression of the ATOH1 transgene under control of supporting cell-specific regulatory elements becomes silenced in regenerating hair cells as they differentiate, and this reduction in transgene expression promotes further maturation towards a more mature type II HC fate.
AAV8 displays robust tropism for vestibular supporting cells after IDPN damage in adult mice in vivo
So far, the ex vivo findings indicated that AAV-ATOH1 under control a supporting cell-specific regulatory element enhances maturation of regenerated hair cells, thereby serving as a promising gene therapy candidate for vestibular hair cell regeneration in vivo. We next assessed the in vivo tropism of various capsids in an established model of hair cell damage.
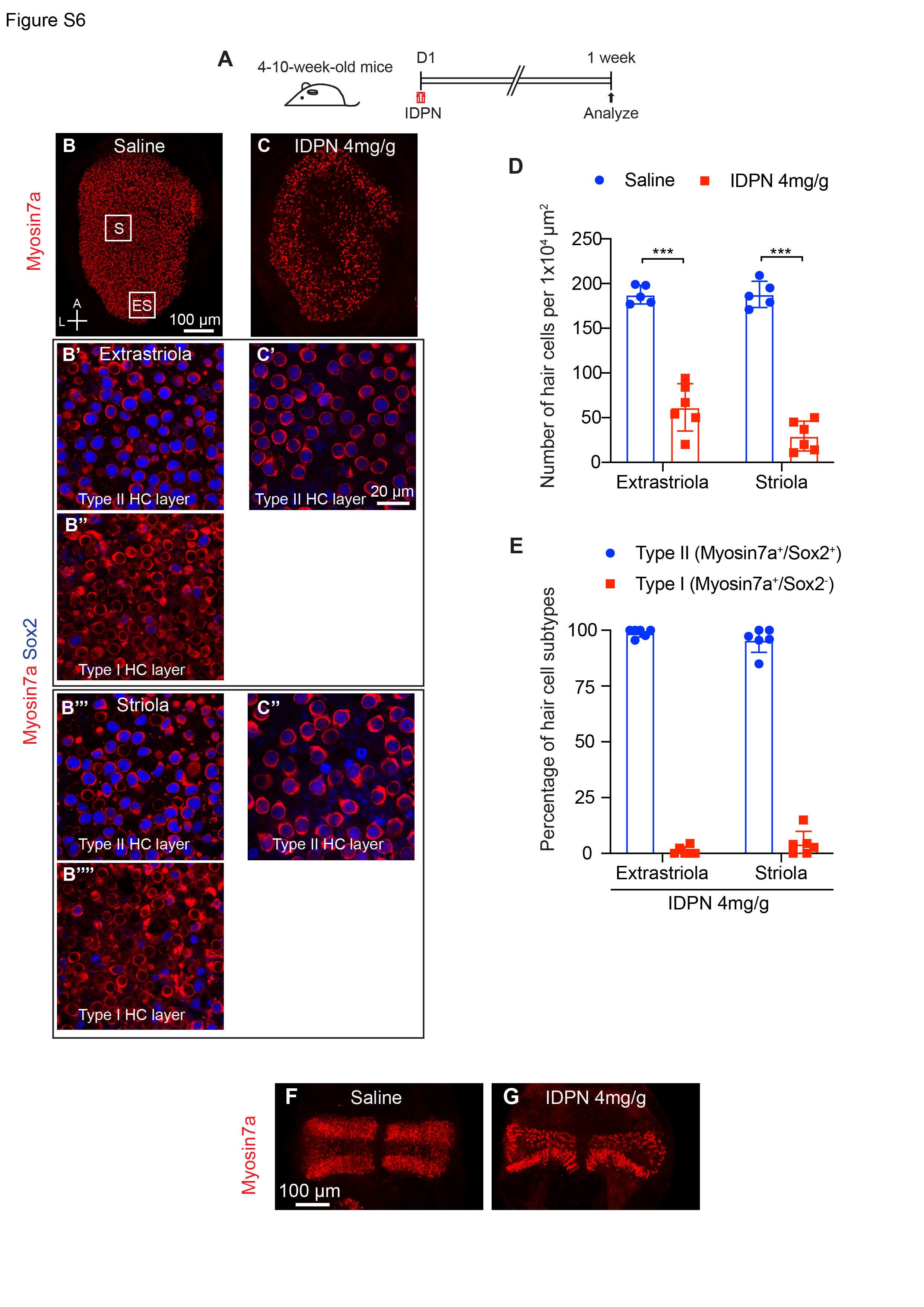
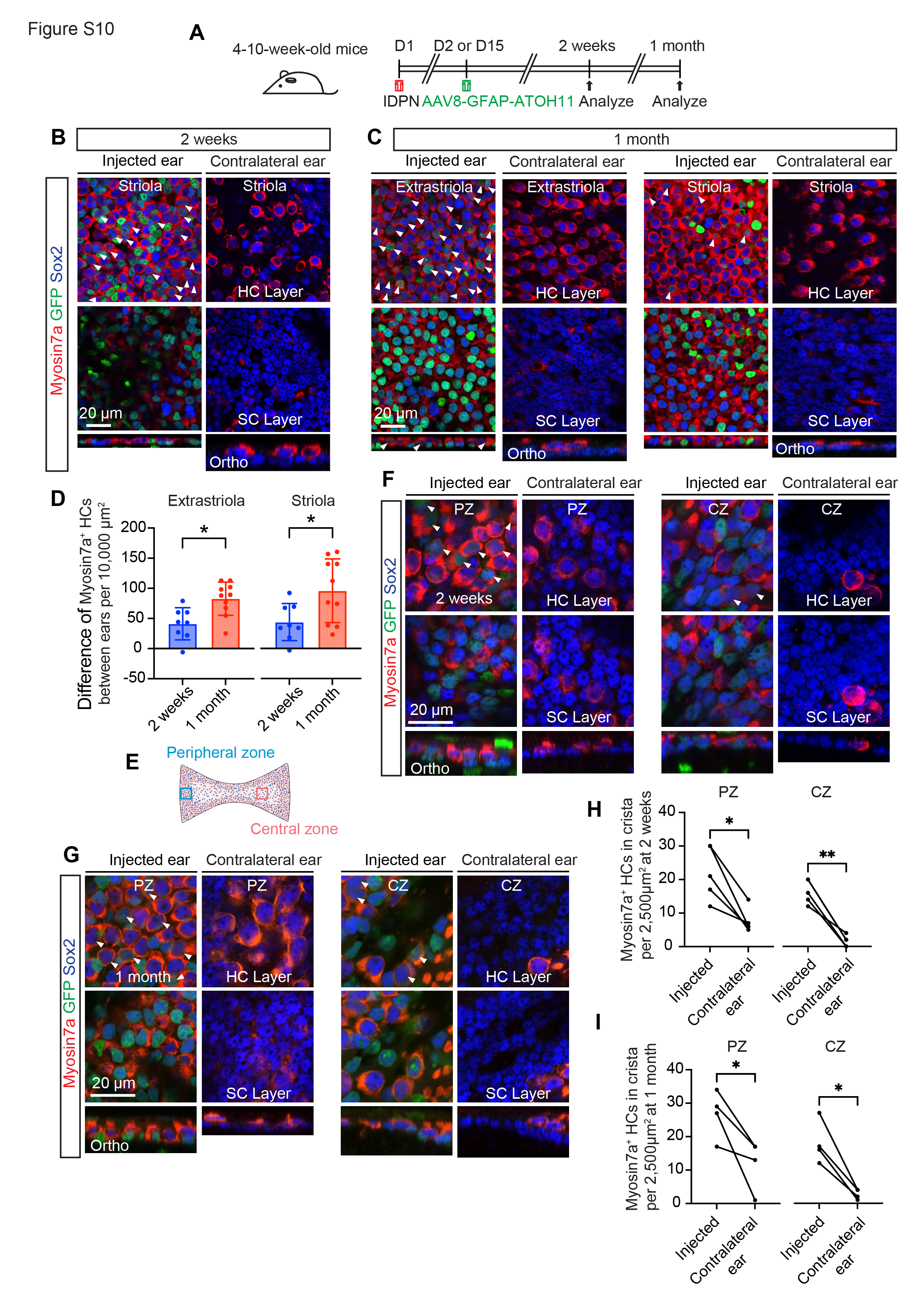
3,3’-iminodipropionitrile (IDPN) is a well-described vestibulotoxin that induces dose-dependent HC death after a single systemic administration to adult mice in vivo13,50. In 4–10-week-old mice, IDPN administration resulted in significant loss of HCs in utricular maculae (67% loss in extrastriola, 84% loss in striola, respectively, p < 0.001) and cristae (61% loss in peripheral zone, 93% loss in central zone, respectively, p < 0.01) within one week (Fig. S6A-G). Hair cell loss was comparable between the striolar and extrastriolar regions of utricles, and the lesions across both ears were highly symmetric in each mouse (Fig. S6A-D). Immunolabeling with antibodies to the type II HC marker Sox2 demonstrated that IDPN preferentially killed type I HCs as 96% of the surviving HCs were Sox2+ (Fig. S6E).
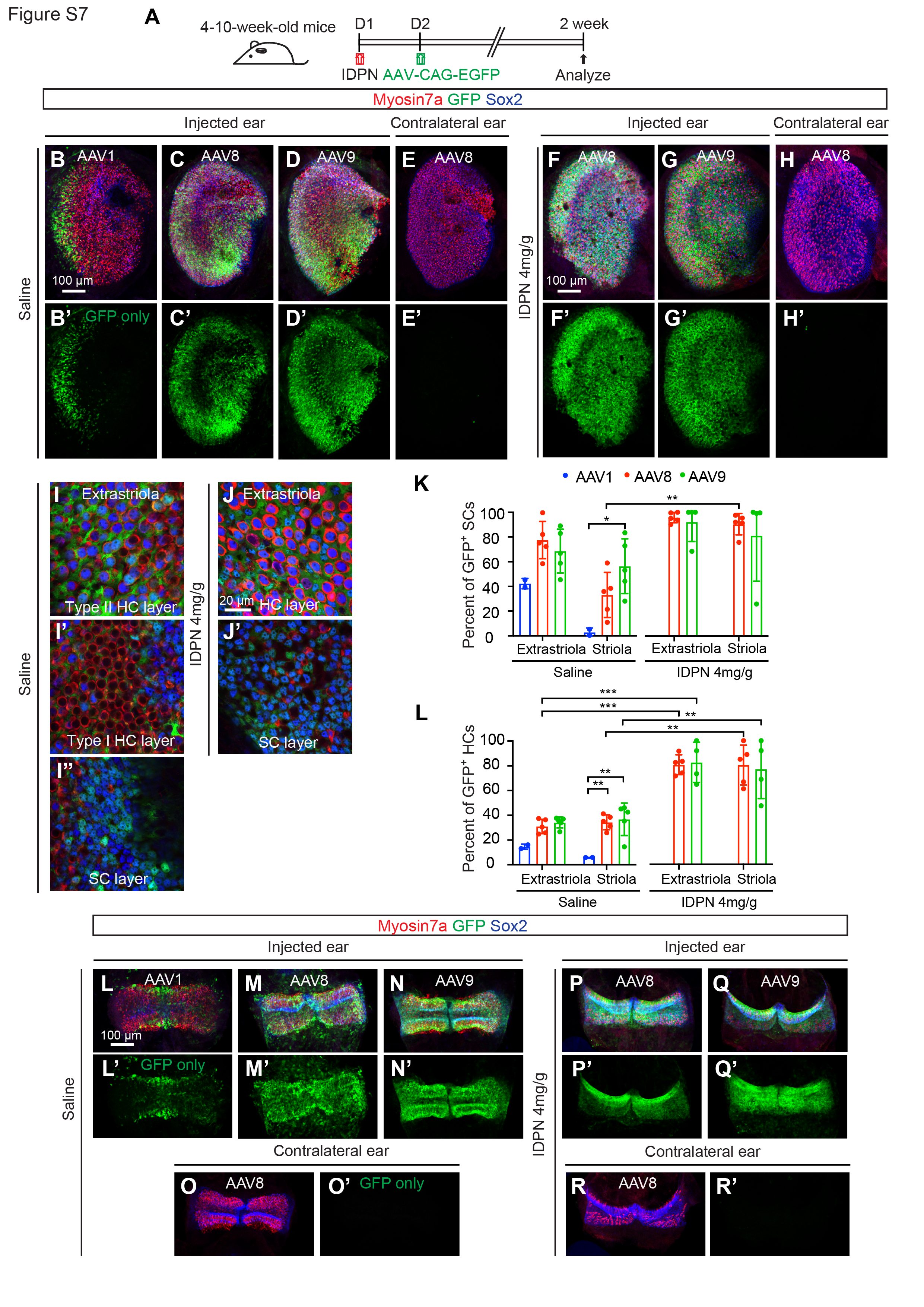
To identify a capsid with optimal supporting cell tropism in IDPN-damaged vestibular organs in vivo, AAV1-, AAV8-, or AAV9-CAG-H2B-EGFP was administered via the posterior semicircular canal (PSC) one day after treatment with saline or IDPN. Two weeks later, vestibular organs were harvested and examined (Fig. S7A). Analysis of the resulting EGFP expression revealed that supporting cell transduction with AAV8 and AAV9 in utricular maculae and cristae was more robust than AAV1 in both the saline control and IDPN conditions (Fig. S7B-D, L-N). Furthermore, the transduction with AAV8 and AAV9 in utricular supporting cells and hair cells appeared higher after IDPN damage, reaching > 78% transduction rates for both cell types (Fig. S7F-G, K-L). On average, AAV8 transduced slightly more supporting cells than AAV9 in the IDPN-treated utricle, with > 90% of supporting cells expressing EGFP in the extrastriola and striola. Of note, no EGFP expression in the contralateral ear was observed in any condition (Fig. S7E’, H’, O’, and R’), indicating that the contralateral ear could serve as untreated controls for ATOH1 gene therapy.
AAV8-GFAP-ATOH1 drives supporting cell-specific transgene expression in adult mouse vestibular organs in vivo
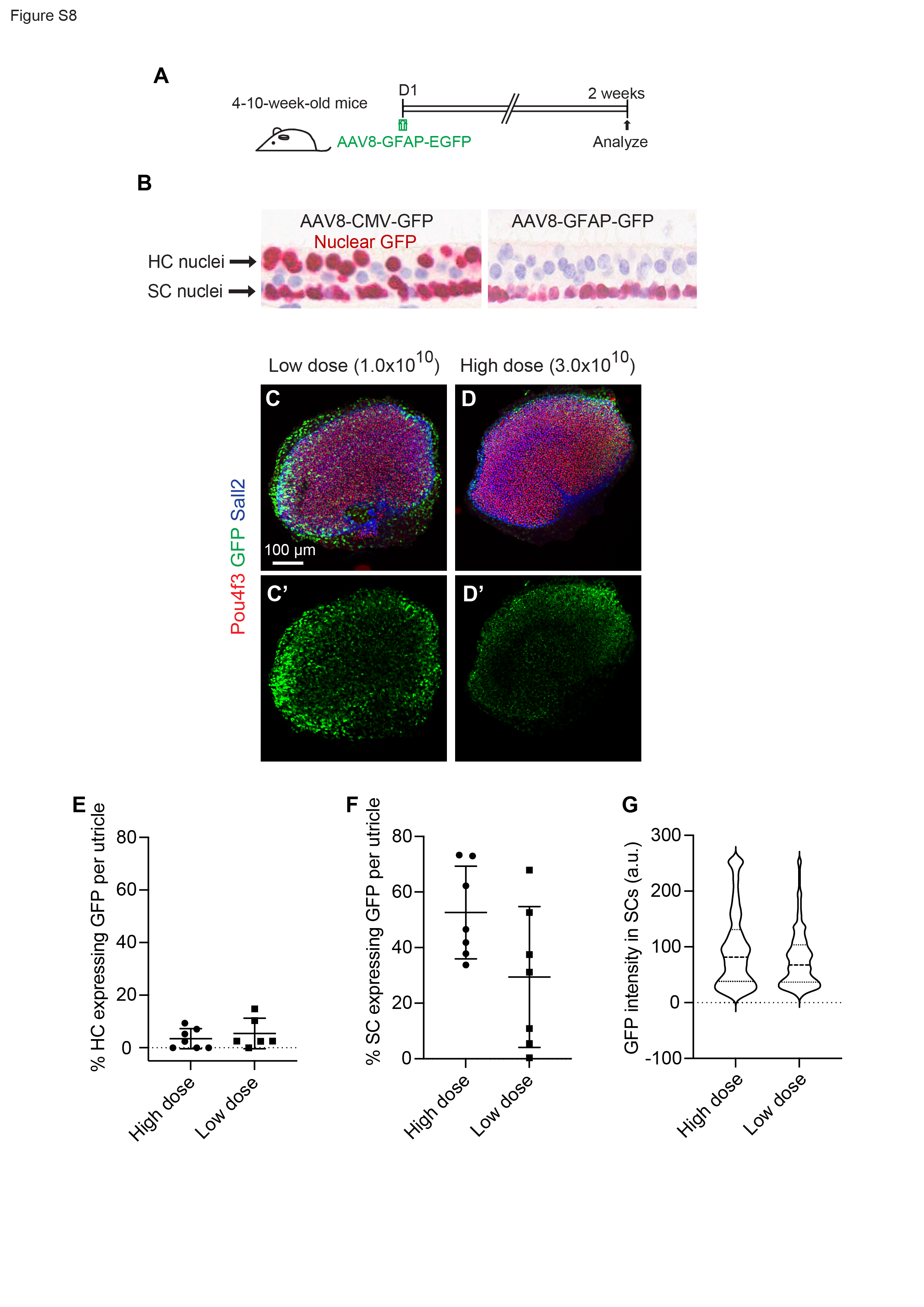
Amongst the three regulatory elements tested ex vivo, GFAP was deemed optimal because it led to strong and targeted ATOH1 expression in supporting cells. To further assess the potential utility of GFAP as a regulatory element for ATOH1 gene therapy, its activity profile was assessed in adult mouse utricle and cristae in vivo. Based on the above results, AAV8 was selected as the capsid and AAV8-GFAP-H2B-EGFP was administered locally at two doses (1x1010 and 3x1010 vg/mL per ear) via the PSC of adult mice (Fig. S8A). Two weeks later utricles showed robust EGFP in supporting cells, with little to no detectable EGFP expression in hair cells (Fig. S8B). No statistically significant difference was observed between the two doses (Fig. S8C-G). These results indicate that GFAP drove robust EGFP selectively in supporting cells in vivo.
AAV8-GFAP-ATOH1 induces regeneration of vestibular HCs with stereocilia bundles and presynaptic ribbons in vivo
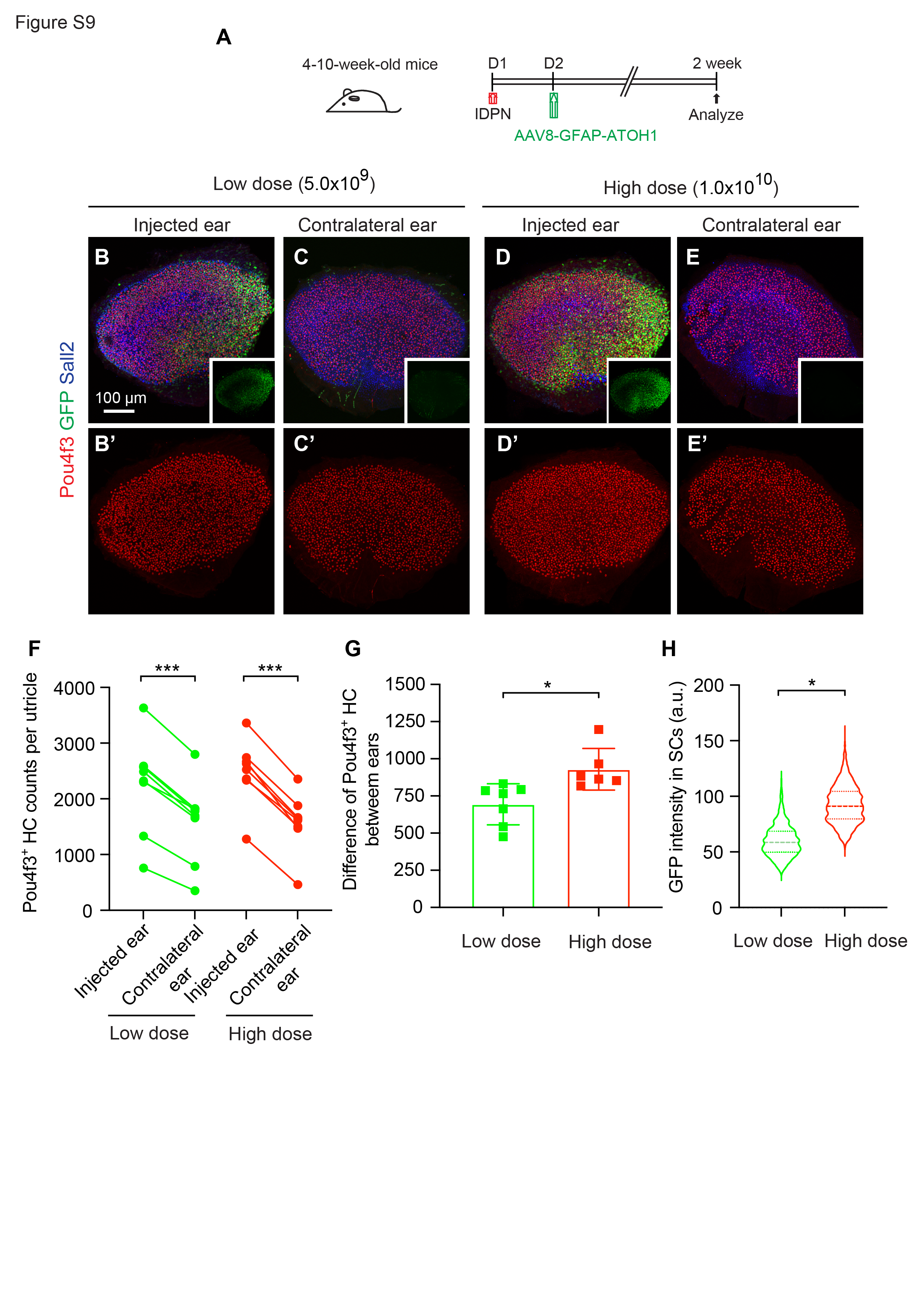
To assess whether AAV8-GFAP-ATOH1-H2B-EGFP promotes regeneration of vestibular hair cells in IDPN-treated mice in vivo, the virus was administered unilaterally one or 14 days after IDPN treatment. Vestibular organs were harvested and examined 2–4 weeks after AAV administration (Fig. 4A). Virus injection resulted in robust expression of EGFP in both utricles and cristae, whereas no EGFP was detected in uninjected contralateral ears (Fig. 4B-D’, G-H, insets). Utricles and cristae from injected ears also displayed more Myosin7a + HCs in comparison to contralateral controls (Fig. 4B-D’, G-H’’, S10A-C, E-H). Quantification showed significantly more Myosin7a + HCs in both striolar and extrastriolar regions of utricles from injected ears compared to contralateral controls at 2 weeks (70.6 ± 51.5 vs. 26.9 ± 28.0 and 109.7 ± 62.3 vs. 68.6 ± 43.8 cells per 10,000 µm2 in striola and extrastriola, respectively) and 1 month (142.7 ± 83.5 vs. 47.0 ± 36.7 and 176.4 ± 50.8 vs. 93.7 ± 36.2 cells per 10,000 µm2 in striola and extrastriola, respectively) (Fig. 4B-C’’, E and F). Similarly, significantly more regenerated hair cells were observed in the peripheral and central regions of cristae 2 weeks (22.0 ± 8.0 vs. 7.6 ± 3.6 and 16.4 ± 3.6 vs. 1.6 ± 1.7 cells per 10,000 µm2 in peripheral and central regions, respectively) and 1 month after AAV administration (26.8 ± 7.1 vs. 12.0 ± 7.6 and 18.0 ± 6.4 vs. 2.8 ± 1.5 cells per 10,000 µm2 in peripheral and central regions, respectively) (Fig. 4G-H, S10G-I), suggesting that AAV8-GFAP-ATOH1-H2B-EGFP promoted regeneration of vestibular hair cells. Quantification of Myosin7a + hair cells 2 weeks after AAV injection confirmed a significant, dose-dependent, increase in the number of hair cells in both striolar and extrastriolar regions of injected ears compared to contralateral control ears (Fig. S9A-F). At the high dose, 912.4 ± 144.7 new HCs were added to the utricle (Fig. S9G), suggesting that about 57.9% of the degenerated HCs had been replaced. In addition, the numbers of Myosin7a + HCs increased with time in both the striolar (70.6 ± 51.5 vs. 142.7 ± 83.5 cells per 10,000 µm2 at 2 weeks and 1 month, respectively) and extrastriolar (109.7 ± 62.3 vs. 176.4 ± 50.8 cells 10,000 µm2 at 2 weeks and 1 month, respectively) regions of the utricle (Fig. 4E-F, S10D).
Hair cells require stereociliary bundles for mechanoreception and synapses to relay information centrally via afferent neurons51,52. One month after AAV8-GFAP-ATOH1-H2B-EGFP administration to IDPN-treated mice, the density of phalloidin-labeled bundles throughout the utricle of injected ears were 1.8-fold higher than contralateral control ears (p < 0.05; Fig. 4I-K).
Ribbon synapses form along the basolateral surface of vestibular hair cells and are identifiable with the presynaptic marker Ctbp253. Surprisingly, utricular HCs in uninjected ears from IDPN-treated mice showed a significant increase in the density of Ctbp2-labeled puncta relative to HCs from naive, undamaged mice (Fig. 4L-O). This phenomenon has been reported in surviving hair cells before54, but whether it resulted from damage-induced fragmentation of pre-existing synapses, an additive process, or remodeling was unclear. In contrast, utricular HCs from ears administered AAV8-GFAP-ATOH1-H2B-EGFP showed comparable Ctpb2 puncta density relative to naive HCs (Fig. 4O). Among 144 hair cells examined in the virus-treated utricle (n = 6), all of them displayed Ctbp2 + puncta, suggesting that they expressed components needed for synaptic transmission.
AAV8-GFAP-ATOH1 induces regeneration of mature type II HCs in vivo
We further characterized in vivo vestibular hair regeneration after AAV8-GFAP-ATOH1-H2B-EGFP using scRNA-seq. Utricles and cristae were collected from IDPN-damaged mice treated with AAV8-GFAP-ATOH1-H2B-EGFP (IDPN and GFAP-ATOH1), IDPN-damaged mice (IDPN), or naive mice (Control; Fig. 5A, S13A) at 2 weeks and 4 weeks post-treatment (4 weeks and 6 weeks after IDPN administration). The organs were pooled by group, single cells were isolated and processed for scRNA-seq. A total of 12,130; 11,111; and 15,146 cells were collected for the control, IDPN, and IDPN and GFAP-ATOH1 treated utricles, respectively, as well as 18,102; 27,072; and 32,927 cells for the cristae (from 6 mice). Marker gene identification in unbiased clusters of the merged groups revealed rich datasets that included all the various cell types that constitute the utricle and crista: hair cells, supporting cells, transitional epithelial cells (TECs), dark cells, melanocytes, mesenchymal cells, glia, endothelial cells, smooth muscle cells, and immune cells (Fig. S11E-I and S14E-I). In the GFAP-ATOH1-treated utricle and crista, ATOH1 transgene was detected in mesenchymal cells, TECs, supporting cells, and supporting cells presumably converting into hair cells (Fig. S11J and S14J). GFAP regulatory element activity in mesenchyme was expected as GFAP expression has been reported55.
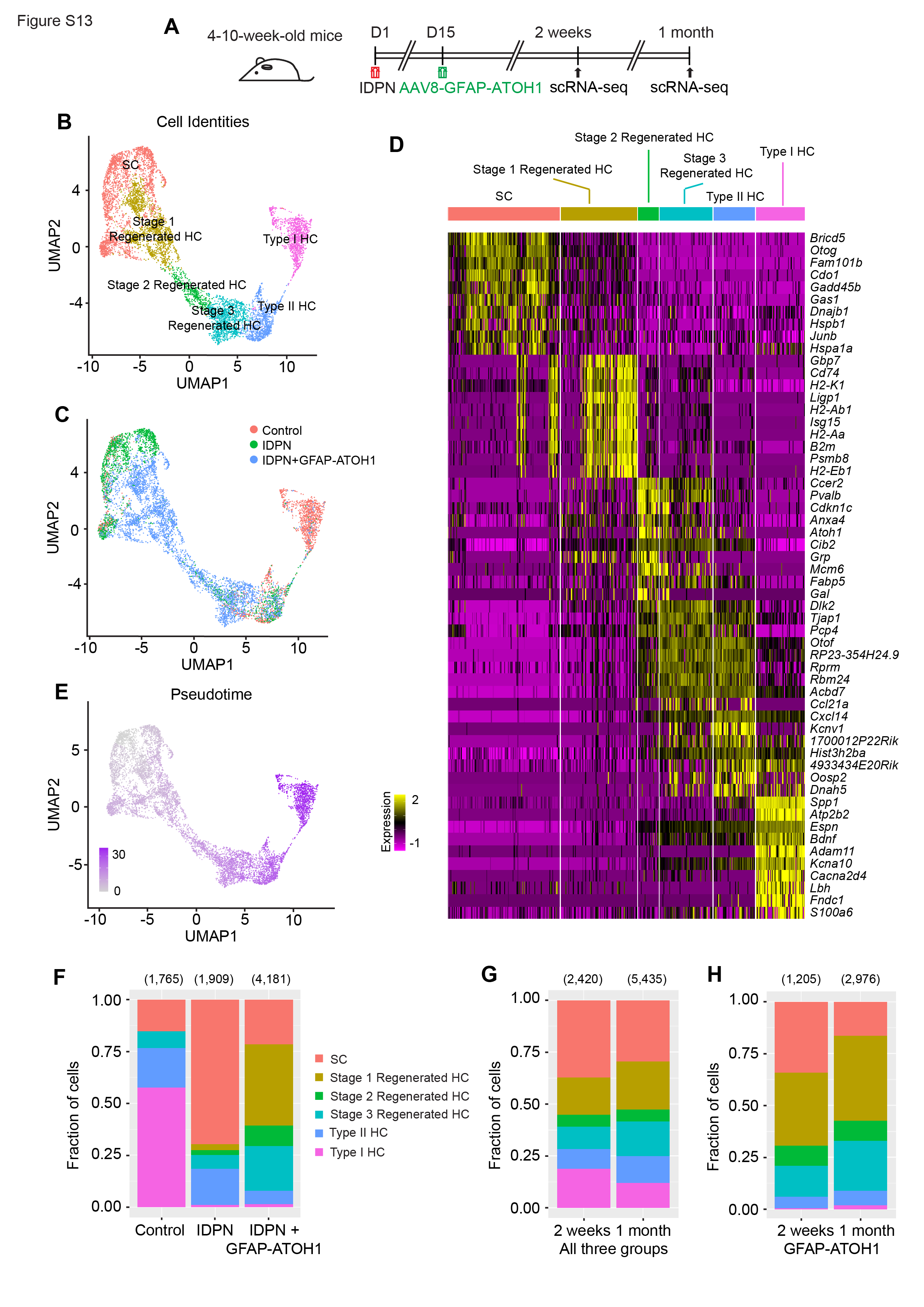
To examine supporting cells converting into hair cells, supporting cell and hair cell transcriptomes from all groups were examined and re-clustered (8,399 cells from the utricles and 7,855 cells from the cristae). Unbiased clustering identified six clusters in both the utricle and cristae (Fig. 5B, D and S13B, D). Marker gene expression revealed three of the clusters were supporting cells, mature type I and II hair cells. Supporting cells and type II hair cells were present in similar proportions for all three treatment groups (Fig. 5F, S13F). In contrast, type I hair cells were scant in the IDPN only and IDPN and GFAP-ATOH1 groups, consistent with the finding that IDPN primarily ablates type I hair cells (Fig. 5F, S6E and S13F)56.
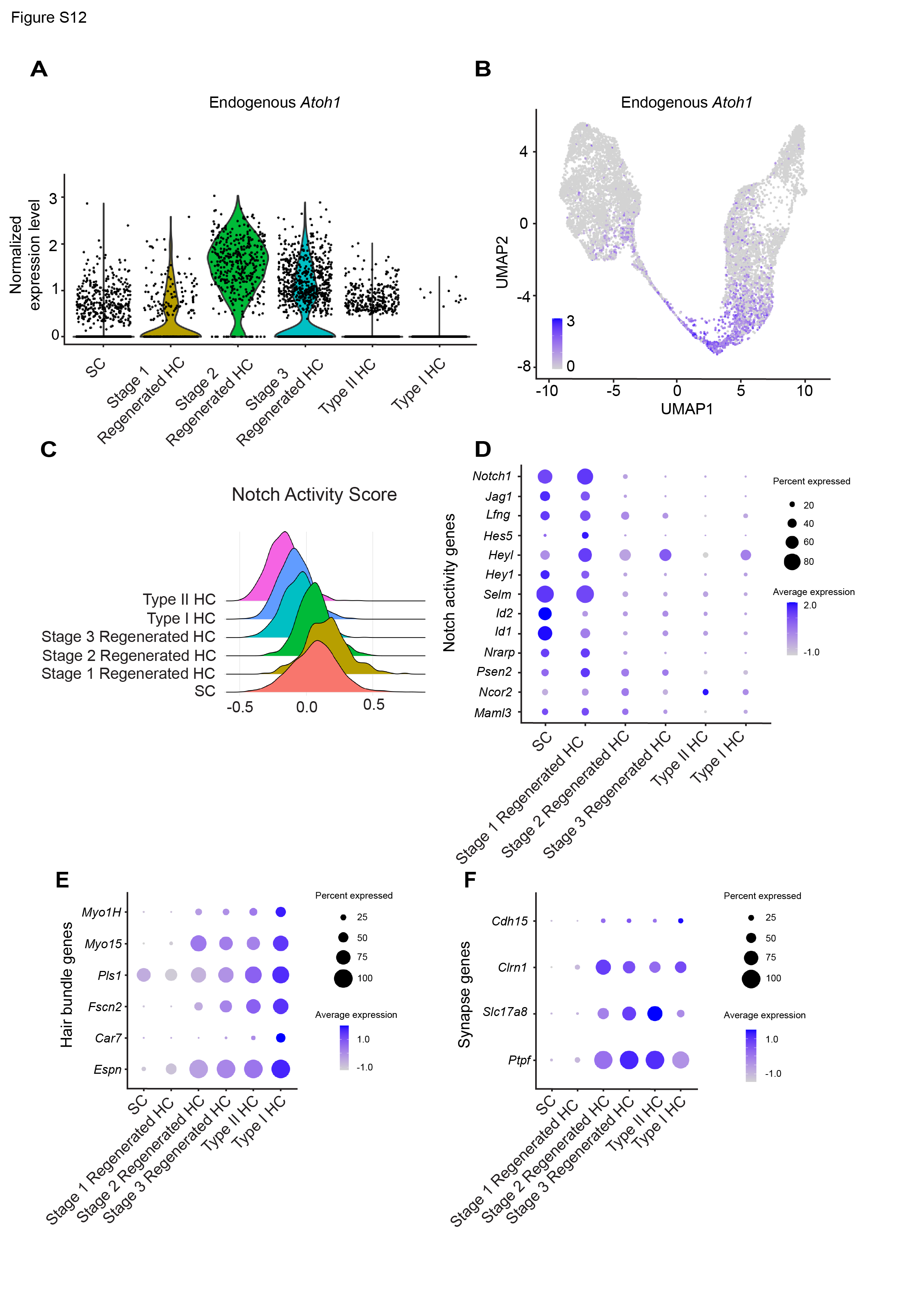
The other three clusters in the utricle primarily comprised cells from the IDPN and IDPN + GFAP-ATOH1 groups. Gene expression analysis indicated that these groups contained cells representing various stages of supporting cell-to-hair cell conversion, thereby revealing the transcriptional map along the supporting cell-to-hair cell axis after in vivo treatment with GFAP-ATOH1. The cells were then ordered along a pseudotime trajectory and were designated stage 1–3 regenerating hair cells, with stage 1 being the earliest stage of conversion and stage 3 being the most mature state (Fig. 5B and E). Out of Stage 1–3 regenerated hair cells, 86.2% came from utricles treated with IDPN + GFAP-ATOH1 (Fig. 5F). These clusters were characterized by hair cell genes known to be expressed during native development and were relatively more mature than converting supporting cells previously described ex vivo (Fig. 5D, Table S4). Compared to undamaged controls, there was an increase in the proportion of stage 1–3 hair cells in the IDPN only group, consistent with a low level of spontaneous regeneration after IDPN damage alone13. Further, there were substantially more regenerating hair cells with ATOH1 overexpression, than after IDPN alone (Fig. 5F). Similar to the ex vivo results, ATOH1 transgene expression levels were highest in supporting cells and stage 1 regenerated hair cells, and then were gradually downregulated in stage 2 and 3 regenerated hair cells (Fig. 6A-B). Thus, like ex vivo, the GFAP regulatory element also became silenced as supporting cells convert into hair cells in vivo (Fig. 6A-B). As the ATOH1 transgene became downregulated, the transcriptomes of the stage 1–3 regenerated HCs scored higher in maturity indices, suggesting a more mature phenotype (Fig. 6C-D, G-H). In addition, based on type I and type II HC indices, regenerating hair cells primarily adopted a type II fate as they progressed from stage 1 to 3 (Fig. 6E-F). No expression of type I hair cell marker genes or increase in type I index score was detected as regenerating HCs differentiated, indicating that ATOH1 overexpression mainly induces a type II hair cell fate in vivo (Fig. 6E-F).
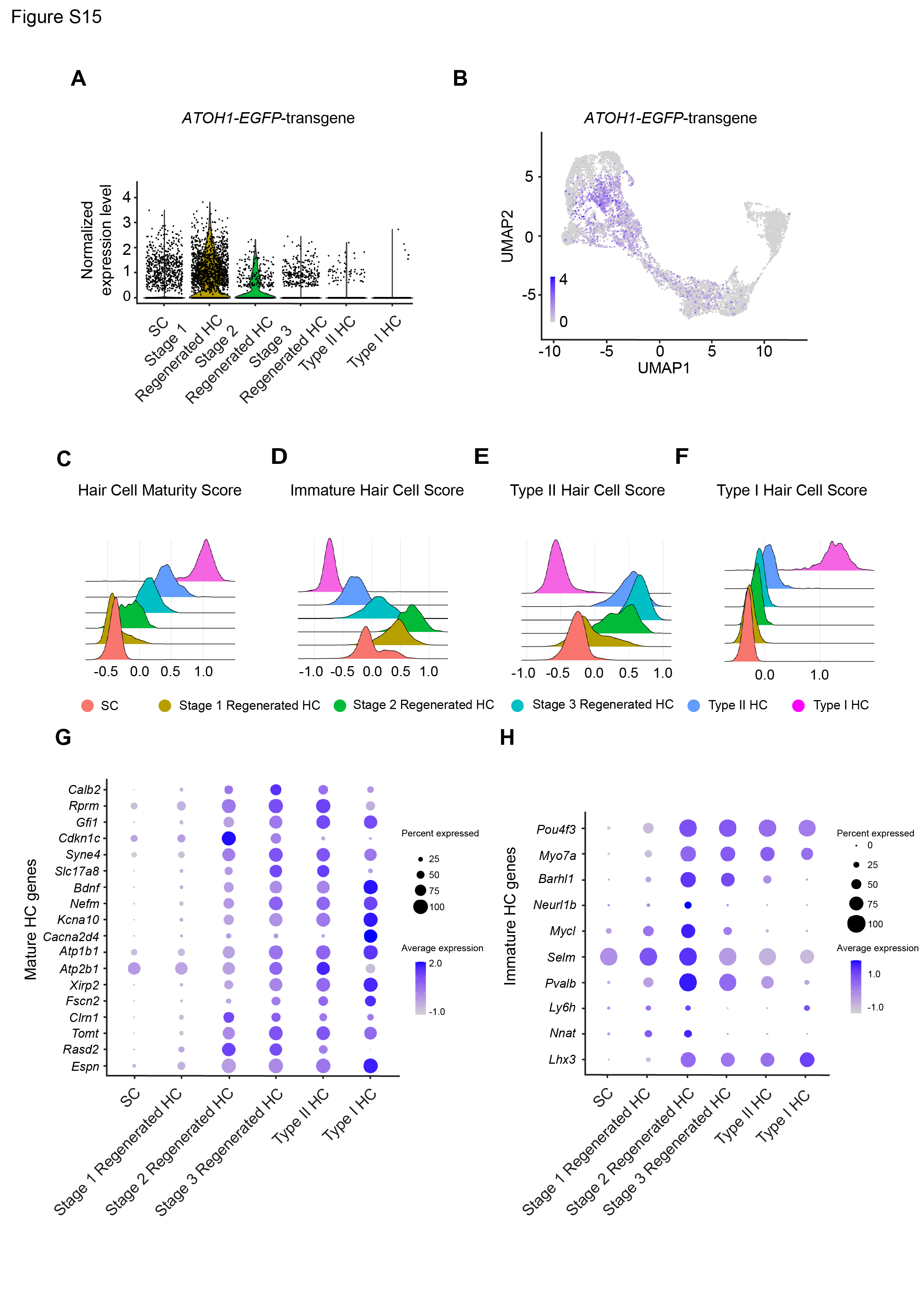
Analysis of the 7,855 supporting cells and hair cells isolated from untreated, IDPN, and IDPN and ATOH1-treated cristae revealed six clusters that are comparable to those found in the utricle (Fig. S13 and S14). In cristae, ATOH1 transgene expression decreased along the trajectory from supporting cells to hair cells (Fig. S15A-B), and hair cells maturity score increased (Figures S15C-D, G-H). At the transcriptome level, newly regenerated hair cells in the cristae also resemble type II hair cells more than type I (Fig. S15E-F).
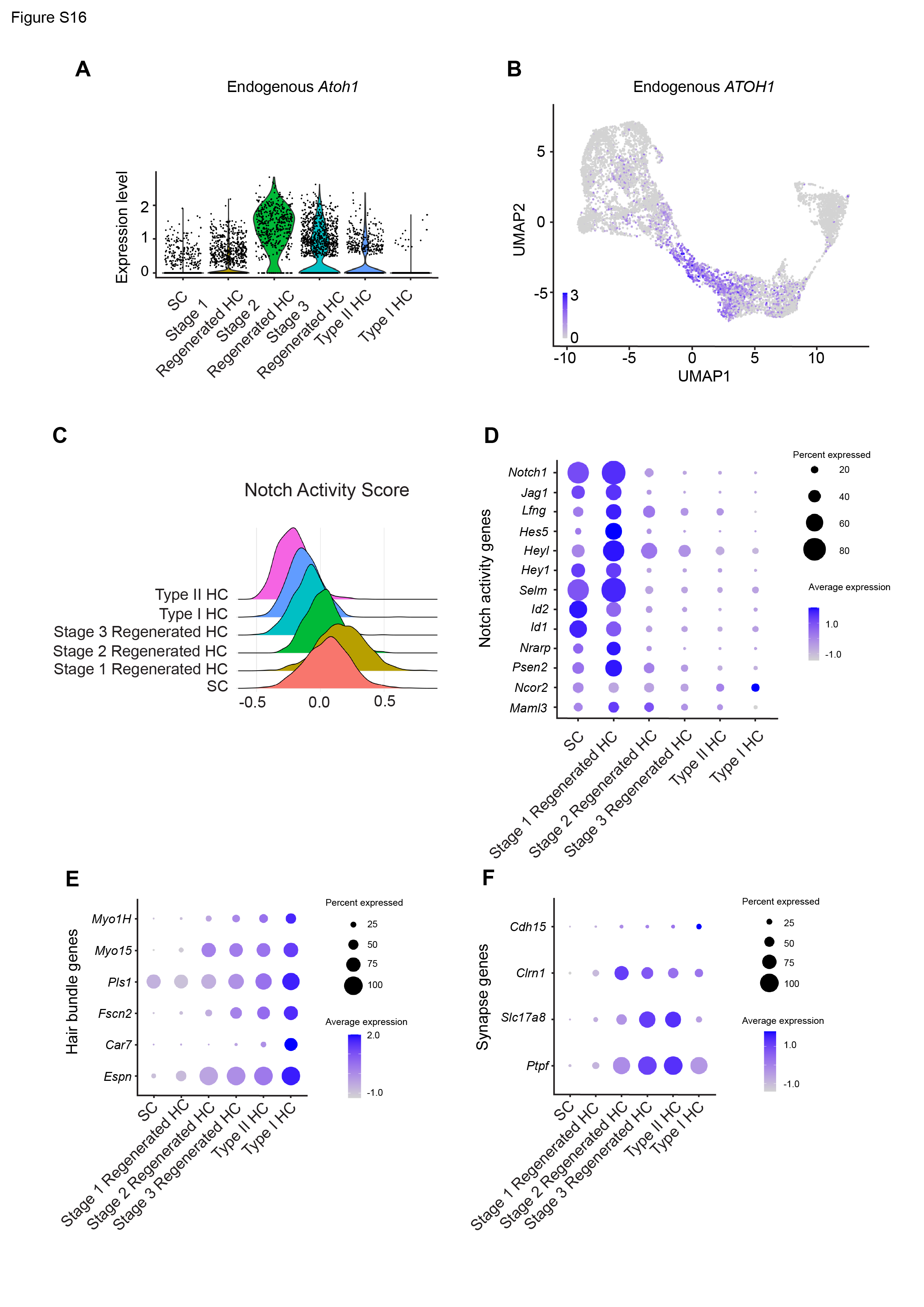
In both the utricle and crista, the expression of Notch pathway genes is highest in supporting cells and stage 1 regenerated HCs and is decreased as maturity increases (Fig. S12C-D and S16C-D). This agrees with the ex vivo results, where Notch activity decreases as hair cells mature.
In summary, these data demonstrate that use of the supporting cell-specific promoter GFAP to drive transient ATOH1 overexpression is effective in enhancing maturation of the regenerated type II-like HCs.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}