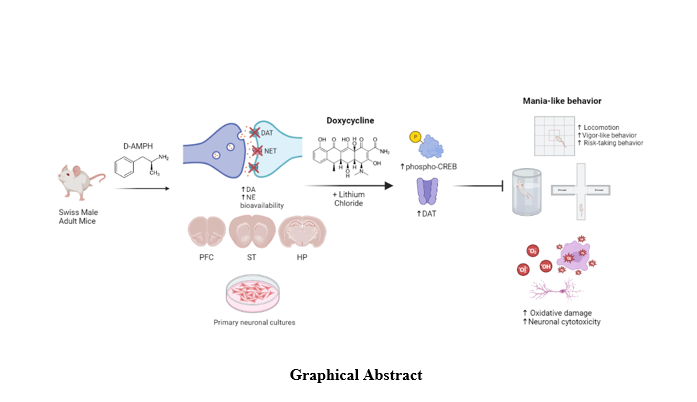
In the present study, we observed that DOXY presents an antimanic-like effect in mice, showing its ability to attenuate behavioral alterations related to hyperlocomotion, risk-taking behavior, and vigor (energy). Also, our results revealed that DOXY protected against the hyperdopaminergic state induced by AMPH and normalized DA metabolite amounts and its turnover rates mainly in the hippocampus, while DOXY+Li also reversed the alterations in PFC DA turnover. In primary cultures, DOXY protected against AMPH-induced cytotoxicity and oxidative damage to hippocampal neurons. Together, our data demonstrated that DOXY attenuated most of the behavioral and neurochemical changes induced by AMPH, an effect possibly associated with a regulation of the hyperdopaminergic state induced by this model.
Regarding mania models, psychostimulants-induced manic-like symptoms have been consistently used to predict new antimanic agents (Macedo et al. 2013; Valvassori et al. 2019a; Chaves Filho et al. 2020a). The most studied of these models is the AMPH-induced mania model. Hyperactivity is a well-characterized aspect of mania and has been traditionally used as the main measurable feature in this model (Frey et al. 2006; Macedo et al. 2013; Valvassori et al. 2019a). Here, as expected, AMPH induced a typical hyperlocomotion in mice, as demonstrated by the increase in zone transition number and exploratory behavior in the OFT, being this alteration reversed by DOXY alone and combined with Li.
In rodents, anxiety, and risk assessment can be evaluated by the exposition to an aversive environment (Swann et al. 2005; Izci et al. 2016). In the EPM test, increased time spent and exploration in the open arms of the maze is recognized as an index of risk-taking behavior in mice and the opposite as anxiety-like behavior (Rodgers and Dalvi 1997; Borta and Schwarting 2005). It was previously demonstrated that AMPH exposure increase premature response and impaired decision making in rodents (Palm et al. 2014; Tremblay and Winstanley 2016). However, in the EPM, there are discrepancies in the literature regarding the AMPH effects, while some studies showed that a single AMPH injection induced an anxiogenic-like response in EPM (Biała and Kruk 2007), this does not seem to be the same for repeated AMPH administration. Of note, Gatica et al., 2017 showed that AMPH sensitization (5 days, daily, 1.5 mg/kg) is not accompanied by anxiety in the EPM test (Gatica et al. 2017). Further, Zhou et al., 2015 reported that repeated AMPH (14 days, daily, 2 mg/kg) induced a risk-taking behavior in EPM and wire-beam bridge tests, and these behavioral changes were attenuated by Li treatment (Zhou et al. 2015).
Here, we showed, for the first time, the ability of DOXY alone or combined with Li to reverse AMPH-induced risk-taking behavior in the EPM. Previously, our research group demonstrated the effect of the correlate tetracycline minocycline (MINO) in the reversal of risk-assessment deficits induced by DA reuptake inhibitor - GBR12909 in mice. MINO, like the observed here with DOXY, reversed risky behavior and hyperlocomotion in the GBR12909-mania model (de Queiroz et al. 2018).
Another domain of mania is increased vigor and goal-directed behavior. In the DSM-V, vigor behavior is included in the clinical diagnosis of BD I and II (Johnson et al. 2012; Rowland and Marwaha 2018). Conceptually, the increased vigor in mania can be perceived as the opposite of anergy and hopelessness diagnosed in depression (Kennedy 2008). The most frequently used animal model for this phenotypic domain is the FST(Detke et al. 1997; Costa et al. 2013). The FST has also been proposed as a useful model to measure vigor behavior in a behavioral battery for BD mania. It was demonstrated that inbred hyperactive mice show reduced immobility and increased vigor-like behavior, and mood stabilizers reversed it (Flaisher-Grinberg and Einat 2009). Here, we showed that AMPH markedly increased swimming time, extending the face validity of this model for BD mania. DOXY combination with Li efficiently restored this parameter to normal levels. Li alone was also efficacious in reversing this behavior.
The comprehension of the biological substrates underpinning BD is very poor. Compelling data has postulated the concept of cycling DA changes in bipolar brains (Berk et al., 2007). Here, AMPH caused a sustained increase in dopamine (DA) levels and potentiated DA neurotransmission. The mechanisms for these AMPH-induced dopaminergic changes include i) inhibition of DA transporter (DAT) by competitive antagonism, ii) stimulation of the vesicular release of DA, and iii) DA reverse-transport towards synaptic cleft independently of the membrane potential (Daberkow et al. 2013; Calipari and Ferris 2013).
Regarding DA metabolism and turnover, here, in the PFC and hippocampus, AMPH caused a reduction in HVA and DOPAC levels, as well as a reduced DA turnover ratio. However, in the striatum, paired with increased DA levels, AMPH markedly augmented HVA levels at the cost of DOPAC reduction. Firstly, it is worth mentioning that few studies directly assessed the levels of monoamines and their metabolites in limbic brain regions in the AMPH mania model. Interestingly, these findings dialogue with previous clinical studies showing elevated levels of HVA and reduced DOPAC levels in CSF of BD patients in a mania state (Swann et al. 1983; Gerner et al. 1984). Increased HVA and DA metabolization was also demonstrated in other rodent models caused by DAT dysfunction, such as DAT knockdown mice and GBR12909 (Milienne-Petiot et al. 2017; Kwiatkowski et al. 2019). Together, these data provide additional construct validity for this animal model. The discrepant findings in the PFC and hippocampus can be attributed to the partial inhibition of MAO and COMT exerted by AMPH in these brain areas, and the subsequent rise in the toxic derivatives of DA metabolism, but this needs to be further clarified.
AMPH also causes abnormalities in other neurotransmission systems. In this regard, noradrenaline transporter (NET) and serotonin transporter (SERT) are important targets for psychostimulants. AMPH binds with high affinity to NET (Ki = 0.07-0.1 µM) and DAT (Ki = 0.1-0.6 µM) and with moderate affinity to SERT (Ki = 20- 40 µM) (Han and Gu 2006). As abovementioned, AMPH is a substrate not only for transporters but also for vesicular monoamine transporters (VMATs). Like DA, AMPH accumulates in synaptic vesicles, changing the proton gradient toward neurotransmitter accumulation in the cytosol (Buchmayer et al., 2013; Steinkellner et al., 2015).
Accordingly, we observed that AMPH induces a marked increase in NE levels in all evaluated brain regions but had mixed actions in serotonergic neurotransmission. Indeed, AMPH increased 5-HIAA levels and potentiated 5-HT turnover, mainly in the hippocampus and striatum. Serotonin is released via dense long-range projections throughout the limbic brain. Despite drugs acting in 5-HT signaling classically being associated with an antidepressant action (Andrews 2015), much less is known about 5-HT neurotransmission in mania neurobiology. An elegant study showed 5-HT depleted Tph2 mutant mice showed a maladaptive behavior resembling BD mania, such as increased risk-taking behavior and aggressiveness. Interestingly, these alterations were reversed by valproic acid treatment (Maddaloni et al. 2018). Therefore, our results are compatible with the increased release of catecholamines (DA and NE) and their subsequent metabolism caused by AMPH and other psychostimulants. Also, the sustained release of monoamines and its reverse flux in the cytosol, caused by AMPH, is a potential factor mediating the increased production of reactive oxygen species (ROS) and other toxic metabolites, which can damage the serotonergic synaptic terminals. However, this needs to be further demonstrated (Berman and Hastings 1999; Dela Peña et al. 2015).
In our experimental conditions, DOXY alone or combined with Li normalized the hyperdopaminergic state caused by AMPH, mainly in the hippocampus. However, compared to Li, DOXY was less efficient in counteracting the depletion of 5-HT levels and the increase in the 5-HT turnover rate induced by AMPH. It was previously demonstrated that DOXY showed important neuroprotective properties in models related to DA imbalance, such as Parkinson's disease (Lazzarini et al. 2013) and schizophrenia (Ben-Azu et al. 2018a). Similarly, MINO significantly attenuated the increase in striatal DA extracellular levels after MDMA administration (Zhang et al., 2006). Therefore, the normalization of DA neurotransmission can be a putative mechanism by which neuroprotective tetracyclines (such as MINO and DOXY) can counteract the abnormal neurotransmission involved in multiple neuropsychiatric conditions.
The cAMP-response element binding protein-CREB is an intracellular protein that regulates the expression of genes that are important in dopaminergic neurons since DA affects the phosphorylation of CREB via G protein-coupled receptors. Still, CREB signaling is a major pathway involved in neurogenesis and synaptic plasticity (Ortega-Martínez 2015). It is well known that mood stabilizers and antidepressant agents increase their phosphorylated (activated)-status (Budziszewska et al. 2000). As expected, in our in vivo protocol, AMPH considerably impaired CREB phosphorylation in the hippocampus. Still, regarding the in vivo protocol, we observed that only Li treatment rescued phospho-CREB expression to a control level, but this result did not reach statistical significance.
Compelling evidence has supported the involvement of abnormal functioning of the dopamine transporter (DAT) in the neurobiology of BD. Single nucleotide polymorphisms (SNPs) in the DAT gene are more prevalent in BD patients than in the general population (Pinsonneault et al. 2011). Conversely, mice with genetic deletion of DAT (DAT-KO) showed marked behavioral alterations resembling mania in cross-translational behavioral tests (Young et al. 2011). Here, AMPH markedly decreased hippocampal DAT expression, which is compatible with the damage to dopaminergic terminal caused by AMPH toxicity. Only Li efficiently restored DAT expression in this brain area.
Pro-oxidative alterations are present in all phases of BD but achieve high levels in manic phase (Kunz et al., 2008). The hyperdopaminergic state can create a pro-oxidative milieu responsible for damaging cell structures. In fact, with DA accumulation, its metabolization by MAO is increased, leading to the formation of the radical hydrogen peroxide (H2O2) and DOPAC (Asanuma et al. 2003). Alternatively, DA can undergo nonenzymatic hydroxylation in the presence of Fe2+ and H2O2, producing 6-hydroxydopamine (6-OHDA), a potent neurotoxin (Graham et al. 1978; Kannari et al. 2006). Considering oxidative stress as a direct marker of DA toxicity, we evaluated the effects of DOXY against AMPH-induced changes in cell viability and oxidative stress markers.
Here, AMPH reduced hippocampal cells’ viability, an effect reversed by DOXY and Li. Also, DOXY alone or combined with Li not only decreased lipid peroxidation but also restored the GSH content of hippocampal neurons exposed to AMPH. Therefore, our findings reinforce DOXY’s ability to protect hippocampal neurons against cell damage mediated by the AMPH-induced hyperdopaminergic state. However, the underlying mechanisms for this action should be further clarified. Also, our findings are in accordance with previous studies showing the antioxidant properties of DOXY in mice models (Ferreira Mello et al. 2013; Ben-Azu et al. 2018b). Despite this, here we showed for the first time DOXY’s in vitro antioxidant action in primary neurons exposed to AMPH.
Differently from the in vivo result, we observed that the combination of Li+DOXY effectively reverses the alterations in phospho-CREB expression in hippocampal cells exposed to AMPH. As abovementioned, the activation of CREB is a relevant pathway to mounting the cytoprotective response of mood stabilizers (Jope 2003). Previous reports already showed Li’s ability in protecting rat primary hippocampal neurons against 6-OHDA and MPTP (1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine) induced cell death, through the activation of CREB pathway, and the subsequent increase in the activity of endogenous antioxidant enzymes (Arraf et al. 2012). Our results corroborated these previous findings while also showed a superior effect of Li+DOXY in increasing the phosphorylated CREB isoform. Together, the activation of CREB pathway can represent a putative mechanism for the protection mediated by the DOXY+Li combination against the AMPH-induced oxidative damage in hippocampal neurons.
Limitations and perspectives
The present study had some limitations. First, no existing animal model can completely mimic the human expression of mania. Second, we did not evaluate firing rate of serotonergic or dopaminergic neurons after tested treatments, as well as the binding profile of DOXY in monoaminergic receptors, which can mediate part of the profile of neurochemical changes observed here. Therefore, future studies addressing the expression of D1- and D2-like dopaminergic receptors and the binding affinity of DOXY for these receptors in this animal model of mania may advance the knowledge on the regulation of dopaminergic neurotransmission by DOXY.
{kind=link}