TRIM45 expression is upregulated in bv2 and hippocampus after lps.
In order to comprehensively evaluate the difference of TRIM45 in physiology and septic encephalopathy, we cited the single cell online website (https://singlecell.broadinstitute.org/single_cell) to observe the expression of TRIM45 in normal mouse brain cells (Fig. 1.c), specifically we analyzed three types of cells : neurons, microglia and astrocytes. As shown in the normal situation, the expression of TRIM45 is highest in neurons, followed by astrocytes, followed by microglia. In order to simulate the difference of TRIM45 between septic encephalopathy and normal group, we used lps + ATP to stimulate HT22,BV2,MA cell line, and accidentally found that TRIM45 was significantly expressed in BV2 after modeling (Fig. 1.d). In addition, compared with the control group,the expression of TRIM45 was increased in BV2 after modeling by western blot and immunofuorescence (Fig. 1.e,f). The disturbance of consciousness caused by septic encephalopathy is mainly distributed in the CA1,CA3 and DG regions, so we again cited the above single-cell online website to analyze the changes of TRIM45 in different regions of the hippocampus of normal mice.The result showed TRIM45 evenly distributed in the CA1,CA2,CA3 and DG regions (Fig. 1.g). In order to detect the changes of TRIM45 in different regions of hippocampus of septic encephalopathy, we found that the number and intensity of TRIM45 fluorescence was increased in different regions of hippocampal tissue of septic mice compared with normal mice (Fig. 1.h). In addition, we used immunofluorescence co-localization and double staining of TRIM45 and IBa-1,the result showed TRIM45 was colocalized with Iba-1 in the hippocampus of septic mice (Fig. 1.i). western blot and PCR experiments showed that the expression of TRIM45 in hippocampus of septic mice was higher than normal mice (Fig. 1.j,k). In general, these data show that the expression of TRIM45 was significantly up-regulated in bv2 and hippocampus after septic encephalopathy.
TRIM45 promotes NLRP3 signaling activation in BV2 after lps + ATP.
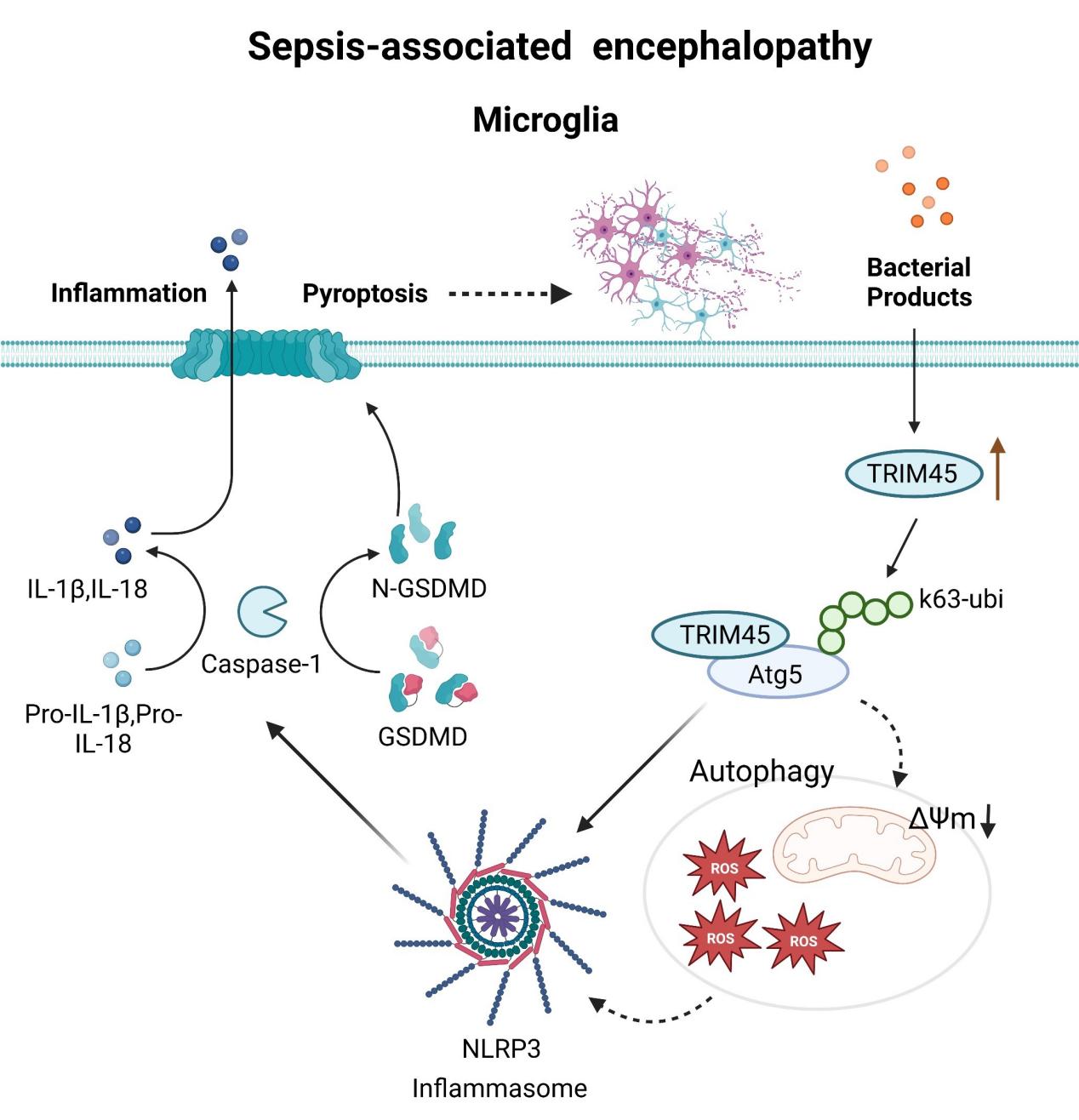
In order to explore which signal pathways are affected by TRIM45, we select the GSE76328 dataset from GEO data, which records the mRNA expression of each gene in U251 cells under the condition of lv-TRIM45 transfection and negative control. Using R-studio software analysis, through the collation and extraction of the two groups of repetitive genes, re-integration and standardization into two groups(C,T); Volcanic map analysis of the integrated data showed that there were 41 up-regulated genes and 30 down-regulated genes (Fig. 2.a); the differential genes were analyzed by KEGG and GSEA, and the results showed that TRIM45 was enriched in the Nod-like signaling pathway (Fig. 2.b,c). NLRP3, a member of the NOD-like receptor (NLRs) family, contains a central nucleotide binding and oligomerization (NACHT) domain, which promotes self-oligomerization and has ATPase activity[18]. There is a close relationship between NLR family and pyroptosis[19]. It has been proved that NLRP3-mediated pyroptosis is one of the pathogenesis of SAE. According to database and literature analysis, we used siTRIM45 to transfect BV2 and studied the relationship between TRIM45 and pyroptosis. First of all, we detected the mRNA levels of IL-1 β and IL-18 in cells and found that under the stimulation of lps + ATP, knockout TRIM45 significantly decreased the levels of these inflammatory factors compared with the model group (Fig. 2.e). In addition, we used Elisa kit to detect the level of IL-1β in extracellular fluid, and also found that knocking out TRIM45 decreased the production of IL-1β(Fig. 2.f). Then we used western blotting to detect the levels of pyroptosis-related proteins in different groups. We were surprised to find that knocking down TRIM45 under the stimulation of lps + ATP decreased the expression of NLRP3 and downstream proteins.(Fig. 2.d). These results were confirmed by immunofluorescence (Fig. 2.h-i). We speculate that the degradation of NLRP3 may be through autophagy rather than proteasome degradation. In addition, we used Hoechst 33342/PI double staining kit to detect cell necrosis by flow cytometry, and found that knocking out TRIM45 can decrease cell necrosis caused by pyroptosis (Fig. 2.g). In summary, these results suggest that TRIM45 can regulate the NLRP3-mediated pyroptosis pathway in BV2.
TRIM45 regulates autophagy proteins associated with NLRP3
It is reported that autophagy-related proteins such as p62, ATG5, and beclin1 play an important role in regulating the activation of inflammatory bodies in NLRP3[20–23]. We try to explore the relationship between TRIM45 and autophagy proteins. We used siTRIM45 to knock down BV2 to explore the mRNA changes of p62, Atg5 and beclin1 under lps + ATP stimulation. The results showed that in the case of modeling, knocking down TRIM45 would decrease the mRNA level of p62, while there was no significant change in beclin1 and Atg5(Fig. 3.a). Similarly, in the case of modeling, knocking down TRIM45 not only decreased the protein expression of p62, but also decreased beclin1, Atg5 and LC3II/I, while overexpressed TRIM45, Atg5, p62, beclin1, LC3II/I increased (Fig. 3.b). We speculate that there may be degradation or modification of posttranscriptional proteins in Atg5 and beclin1. The imbalance of redox homeostasis under pathological conditions will lead to excessive production of ROS. Autophagy is the main cellular defense against oxidative stress or related conditions leading to the accumulation of damaged proteins or organelles[24, 25]. The results show that under the stimulation of lps + ATP, knockout of TRIM45 can significantly reduce the production of reactive oxygen species, and ROS is one of the mechanisms of NLRP3 activation(Fig. 3.e). Mitochondrial membrane potential damage is an important source of ROS production in cells[26]. We continue to study the effect of intervention on TRIM45 on membrane potential. The results showed that under lps + ATP stimulation, knockout of TRIM45 could improve the mitochondrial membrane potential and reduce the mitochondrial membrane damage(Fig. 3.d). According to the above results, under the stimulation of lps + ATP, intervention of TRIM45 will affect autophagy-related proteins, change the change of mitochondrial membrane potential and regulate the production of ROS.
TRIM45 regulates Atg5 based on its own E3 ubiquitin ligase activity
Because TRIM45 is an E3 ubiquitin ligase, it has the function of ubiquitin substrate. In addition, previous studies have shown that TRIM45 can affect the protein level of Atg5, but there is no change in mRNA level. Therefore, we explore whether TRIM45 affects Atg5 protein through its E3 ligase activity. Under the stimulation of lps + ATP, the expression of ATG5 was decreased due to the knocked down of TRIM45 (Fig. 4.a). We used CHX to study the half-life of ATG5 protein in lps + ATP + si-NC group and lps + ATP + siTRIM45 group. The results showed that siTRIM45 accelerated the degradation of ATG5 (Fig. 4.b). We queried the amino acid sequences of TRIM45 and Atg5 through the official website of NCBI, and analyzed the confidence of the two proteins using high-throughput interaction omics. It was found that the score was more than 0.7. There is a possibility of binding between the two proteins. Further co-ip assay in vivo showed that TRIM45 was bound to ATG5 in BV2 (Fig. 4.c,d). In addition, some studies have shown that USP22 can inhibit NLRP3 inflammasome by Atg5 autophagy to degrade NLRP3, in which Atg5 can directly bind to NLRP3. Therefore, we used co-immunoprecipitation to explore whether the intervention of TRIM45 affected the binding of Atg5 and NLRP3. The results showed that compared with the control group, the model group and model + HA-TRIM45 could inhibit the binding of Atg5 and NLRP3(Fig. 4.e). The next step is to explore whether TRIM45 has ubiquitin modification in Atg5 protein. We found that ubiquitin protein in lps + ATP group was increased compared with control group (Fig. 4f). It has been reported that the ubiquitin chain of TRIM45 is mainly K63 and K48. K63 plays a role in stabilizing protein structure, while K48 promotes protein degradation[27, 28]. HEK293T cells were transfected with HA-ub, HA-k63ub or HA-k48ub and TRIM45. The results show that TRIM45 promotes Atg5 ubiquitin through K63 instead of K48 (Fig. 4.g,h,i). Current experiments show that TRIM45 stabilizes its ATG5 stability by coupling ATG5 with K63 ubiquitin chain.
TRIM45 regulated NLRP3 pathway in an Atg5 dependent manner.
It has been reported that Atg family proteins affect NLRP3 pathway by regulating autophagy[29–31]. Previous studies have found that intervention with TRIM45 can change the Atg5 or NLRP3-mediated pyroptosis pathway; TRIM45 promotes ubiquitination of Atg5 with K63. Therefore, we aim to determine whether TRIM45 regulates the NLRP3 pathway in an Atg5 manner. Firstly, Western blotting showed that up-regulation of TRIM45 expression increased the expression of ATG5 and NLRP3 pathway proteins in lps + ATP-stimulated bv2 (Fig. 5.a), while knocking down TRIM45 decreased the expression of ATG5 and NLRP3 pathway proteins (Fig. 5.b). The effect of knocking down TRIM45 on inhibiting NLRP3 activation was reversed by overexpressed Atg5 (Fig. 5.c). In summary, these findings suggest that TRIM45 promotes NLRP3 activation in an Atg5-dependent manner.
Inhibition of TRIM45 can reduce neuronal injury in SAE.
Previous experiments have shown that TRIM45 increased the expression of pro-inflammatory cytokines in BV2 after lps + ATP. In order to further confirm whether TRIM45 knockout can directly protect neurons through microglial regulation in septic encephalopathy, we used siTRIM45 to transfect BV2 to produce a bv2-HT22 co-culture system(Fig. 6.a). TUNEL staining showed that knocking down TRIM45 could save neuronal death caused by lps + ATP stimulation (Fig. 6.c). Western blotting analysis showed that the knockdown of TRIM45, Caspase3 and cl-Caspase3 in lps + ATP was down-regulated compared with the model group(Fig. 6.b). In addition, Nissl staining was used to evaluate lps-induced neuropathy. The results showed that intraperitoneal injection of lps mice showed extensive damage to neurons in hippocampal CA1 and DG regions, while neuronal damage in hippocampal CA1 and DG regions decreased in shTRIM45 mice(Fig. 6.d).
Knockdown of TRIM45 can inhibit pyroptosis, improve cognitive impairment and may be a target for patients with sepsis.
At present, experiments have proved that there is a relationship between TRIM45 and pyroptosis at the cellular level. To further explore the relationship between TRIM45 and pyroptosis in vivo, AAV-Scramble or AAV-shTRIM45 mice were used to carry out immunoblotting and immunofluorescence experiments under lps stimulation. Experiments show that TRIM45 knockdown, such as the results of cell experiments, will inhibit NLRP3 and Gsdmd-N(Fig. 7.a, b). In addition, we explored the effect of intervention of TRIM45 on the seven-day survival rate of septic mice. The results showed that the survival rate of shTRIM45 mice was higher than that of septic mice(Fig. 7.c). At the same time, in order to verify whether TRIM45 is related to cognitive impairment. We conduct MWM tests to explore spatial learning and memory. The results showed that under the condition of lps, the animals pretreated with shTRIM45 had less time to reach the hidden platform, and the TRIM45 knockdown mice spent more time wandering in the targeting quadrant(Fig. 7.d). Based on the above results, we proposed whether TRIM45 can be used as a regulatory protein or marker of sepsis. We screened 47 patients with sepsis, the details of which are shown in Table 2. We extracted peripheral monocytes from patients with sepsis and analyzed TRIM45 by PCR and wb. The results showed that the levels of TRIM45 mRNA and protein in septic group were higher than those in normal group(Fig. 7.e-f). In addition, we analyzed the TRIM45 mRNA level of peripheral blood monocytes and APACHE II score in some patients with sepsis by linear regression analysis, and the results showed that there was a positive linear correlation between them(Fig. 7.g). Therefore, the reduction of brain tissue TRIM45 may alleviate the brain damage caused by sepsis, which is one of the detection targets of sepsis.
Table 2
baseline clinical characteristics of the study subjects.
| Patient group |
| Characteristics | All patients | Survivors | Nonsurvivors |
| Demographics and underlying conditions |
| Number of patients | 47 | 36 | 11 |
| Males,number(%) | 37(78.7%) | 31(86.1%) | 6(54.5%) |
| Age(years) | 73[60.25,81.75] | 70[59,79] | 82[65.5,88.5] |
| Cardiac functional insuffificiency ,number(%) | 10(21.3%) | 8(22.2%) | 2(18.1%) |
| Renal functional insuffificiency ,number(%) | 14(27.8%) | 10(27.8%) | 4(36.4%) |
| Hypertension,number(%) | 27(57.4%) | 18(50%) | 9(81.8%) |
| Diabetes mellitus,number(%) | 24(51.1%) | 17(47.2%) | 7(63.6%) |
| Disease severity,number(%) |
| Septic shock | 23(48.9%) | 14(38.9%) | 9(81.8%) |
| Baseline parameters |
| APACHE II score | 23.11 ± 0.975 | 22.7 ± 1.165 | 24.46 ± 1.686 |
| SOFA score | 8.21 ± 0.515 | 8.23 ± 0.612 | 8.15 ± 0.946 |
| Site of infection,number(%) |
| Lung | 21(44.7%) | 14(38.9%) | 7(63.6%) |
| Abdomen | 20(42.6%) | 16(44.4%) | 4(36.3%) |
| Urinary tract | 3(6.4%) | 3(8.3%) | 0 |
| Other | 3(6.4%) | 3(8.3%) | 0 |
| Intervention,number(%) | | | |
| Mechanical ventilation | 34(723%) | 23(63.9%) | 11(100%) |
| Renal-replacement therapy | 22(46.8%) | 16(44.4%) | 6(54.5%) |
| Length of stay |
| In the ICU(days) | 23.5[12,40.25] | 20[12,33] | 38[16,55] |
| Data is presented as median[interquartile range] or mean ± SEM or n(%). |
{kind=link}