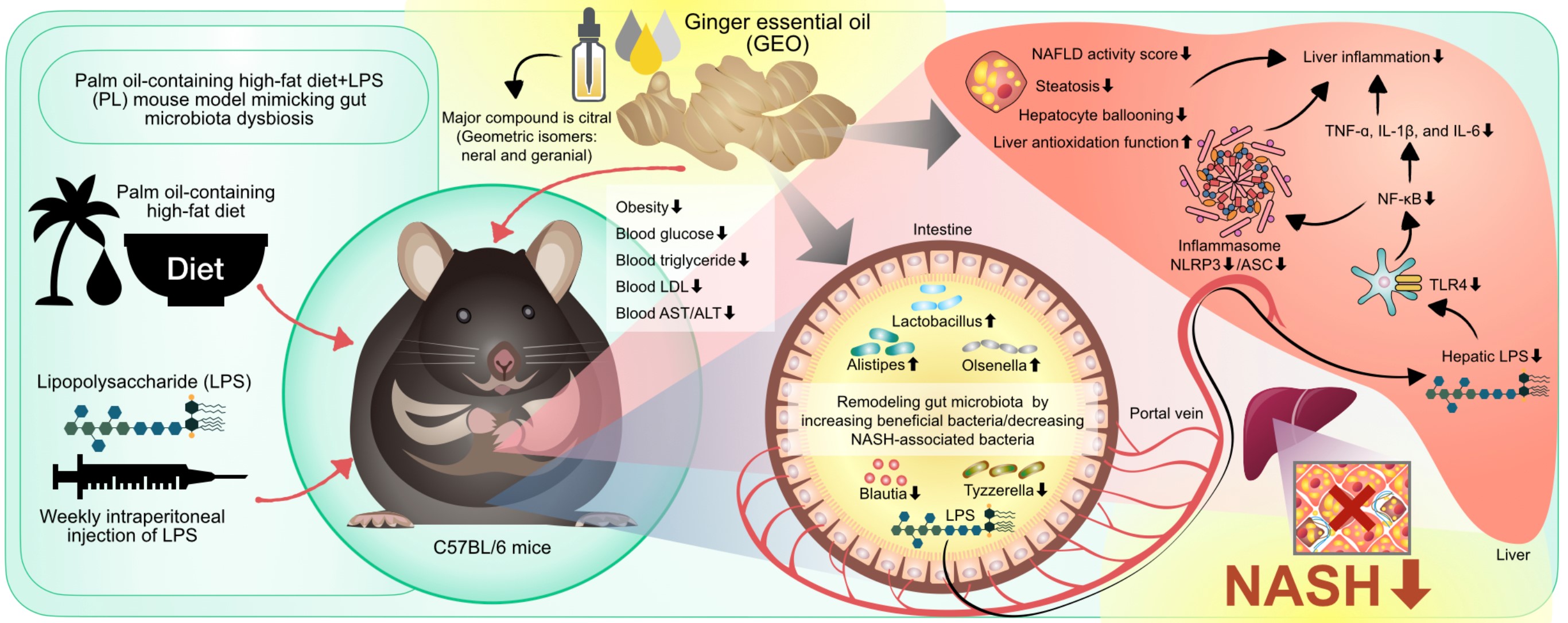
A trans-fat-containing HFD is an excellent model for inducing NASH in animals (24). Due to the prohibition of using trans-fats in the food supply chain and food industry (23), however, trans-fat-containing HFDs may not be suitable for mimicking NASH progression in humans. The replacement of trans-fat with palm oil in a HFD exhibits advantages and the potential for inducing NASH with human NASH phenotype translatability (25). P-HFD-fed rodents manifest morphological features analogous to those of human NASH, including macrosteatosis, lobular inflammation, hepatocyte ballooning degeneration, and periportal/perisinusoidal fibrosis (25). Furthermore, we developed a P-HFD supplemented with an intraperitoneal injection of LPS (PL) to mimic the conditions of gut dysbiosis and endotoxemia. PL causes NASH, intestinal leakage, endotoxemia, and dysbiosis of the intestinal microbiota by increasing pathogenic bacteria and reducing beneficial microbiota (26). Therefore, a PL mouse model was used in this study. Our results demonstrated that PL intervention for 12 weeks negatively affected obesogenic and metabolic biomarkers in mice, including body weight, the area under the curve (AUC) of body weight gain, total fat mass, plasma glucose, total cholesterol, and LDL-C (Fig. 1). The average energy intake of the mice among the groups was not significantly different, suggesting that the adverse obesogenic and metabolic consequences were due to the P-HFDs and LPS injections. The P-HFD diet comprised a large amount of palm oil, fat, fructose, and cholesterol. A previous study found that dietary saturated fat and fructose intake are associated with intrahepatic lipid accumulation, lipogenesis, insulin resistance, oxidative stress, and inflammation (37, 38). Palm oil contains a large amount of saturated fatty palmitic acid (C16:0) (39). Palm oil in normocaloric and normolipidic diets can adversely interfere with and exacerbate metabolic and glucose homeostasis and induce inflammation in the liver and white adipose tissue. The consequence is more acute in mice administered interesterified palm oil (40). Injections of LPS in choline-deficient l-amino-acid-defined (CDAA)-fed mice for 16 weeks have been found to intensify hepatic inflammation and pericellular fibrosis via the TLR4/NF-κB signaling pathway (41). This study demonstrated that the use of PL could be more advantageous than the CDAA diet as PL can mimic the human diet and endotoxemia, whereas the CDAA diet is not practical in humans. In this study, the level of plasma TG declined in the PL group compared to the CON group, which is consistent with previous studies (24, 25) and was likely due to blood cholesterol in the P-HFD inhibiting lipoprotein synthesis, thereby reducing triglyceride secretion from the liver to the circulation (42, 43). In this study, GEO supplementation prevented the obesogenic metabolic phenotype and ameliorated lipidemia by reducing the AUC for body weight and plasma glucose, total triglycerides, LDL-C, and HDL-C. GEO further alleviated the NASH phenotype by attenuating the plasma hepatic damage biomarkers AST, ALT, and NAS, mainly by reducing the hepatocyte ballooning score. Hepatocyte ballooning is generally considered to be a form of apoptosis related to fibrosis and cytoskeletal damage (44). In this study, medium and high doses of GEO (62.5 and 125 mg/kg bw) exhibited a more substantial hepatoprotective effect. GEO displays an anti-hyperlipidemic effect by reducing serum FFA, triglycerides, and total cholesterol levels in mice, previously reported (6). GEO exhibits hepatoprotective properties against alcoholic fatty liver disease and can change particular metabolites in the blood (32).
HFD intake is associated with increased oxidative stress in the liver, which is one of the pathogeneses of NASH (45). NASH patients have been found to exhibit increased CYP2E1, which promotes ROS generation and hepatic oxidative stress (46). Several studies have further demonstrated that ginger exerts antioxidant activity (6, 32, 47). In this study, GEO supplementation substantially lower hepatic CYP2E1 protein expression and significantly enhanced the hepatic antioxidant enzyme capacity, including CAT, GRd, and GSH. These results demonstrate that GEO alleviates hepatic oxidative stress in mice with PL-induced NASH. HFD consumption interferes with homeostasis by impairing lipid metabolism, resulting in imbalanced lipid uptake and storage and leading to hepatic steatosis (7, 48). The PL-induced NASH mouse group showed a substantial increase in the hepatic protein expression of SREBP-1 and HMGCR compared to the control mice. Both of these transcription factors play essential roles in the regulation of hepatic lipid biosynthesis. The elevation of SREBP-1c promotes triglyceride synthesis, and HMGCR is a key enzyme in cholesterol synthesis (49). In addition, PPARα, a regulator of fatty acid β-oxidation, decreased in PL-induced NASH mice. The effect of GEO on these lipid-regulating proteins exhibited a substantial improve, but it did not reach statistical significance, which was consistent with the histopathological data. The different forms of diet-induced NASH models could induce different degrees of NASH severity. PL-induced NASH showed substantial severity compared to our previous study, in which HFD induced simple hepatic steatosis (6, 50).
GEO alleviated the NASH phenotype and pro-inflammatory cytokines TNF-α, IL-6, and IL-1β. Therefore, we investigated the underlying mechanisms of inflammation. The NLRP3 inflammasome plays an essential role in the hepatic inflammatory response (51). NASH patients and animal models have exhibited increases in the activation of the NLRP3 inflammasome (17, 51). Blocking the NLRP3 inflammasome attenuates hepatic steatosis, inflammation, and fibrosis in mice (17). In our study, the expression levels of NLRP3 and ASC were increased in PL-induced mice, whereas they were reduced with GEO supplementation. Caspase-1 protein expression was substantially suppressed by GEO. The activation of the NLRP3 inflammasome is stimulated by danger-associated molecular patterns and PAMPs such as LPS (52). The LPS-mediated TLR4/NF-κB pathway can activate the NLRP3 inflammasome by enhancing the expression of NLRP3 (53). In in vitro experiments, geranial and neral isomers of citral in GEO suppressed the production of the NLRP3 inflammasome and pro-IL-1β (54). Therefore, GEO exhibited anti-inflammatory activity by reducing NLRP3 inflammasome activation, which may be caused by citral and its isomers.
Diet is a primary factor in modifying the composition of gut microbiota (55). PL intervention is a prime factor in modifying the fecal microbiota composition of mice. GEO has less influence on α-diversity. Similarly, PCoA β-diversity based revealed that diet is the principal factor that shapes the gut microbiota. GEO plays a secondary role in modifying gut microbiota bacterial diversity. A high dose of GEO resulted in the gut microbiota shifting outward from the PL-treated group. GEO reduced the relative abundance of NASH-associated bacteria, including Blautia and Tyzzerella, which are considered harmful. Compared to healthy individuals, Blautia abundance increased in NASH patients and was positively correlated with LPS levels (56). Tyzzerella has been reported to be enriched after 12 weeks of HFD intervention and is positively associated with liver dysfunction-associated parameters (57). In contrast, GEO supplementation in PL mice enhanced the relative abundance of Lactobacillus, Alistipes, and Olsenella, which are considered beneficial bacteria. A previous study reported a lower relative abundance of Lactobacillus in GAN ob/ob-NASH mice (24). Lactobacillus may be used as a probiotic to improve steatohepatitis through the gut microbiota–liver axis by modulating gut microbiota composition and the inflammatory pathway in NAFLD (58). Alistipes, which are potential SCFA-producing bacteria that reduce hepatic fibrotic conditions through cytokine modulation, have been found at lower levels in the guts of NASH and NASH-cirrhosis patients (59), (60). Olsenella, an SCFA producer, is associated with tight junction improvement (61, 62). Thus, GEO supplementation can remodel gut microbiota composition and reverse gut microbiota dysbiosis, which may contribute to ameliorating NASH.
The elevated localization of LPS in hepatocytes has been reported in NAFLD and may cause liver inflammation via a TLR4-related pathway (63). This study found no significant changes in intestinal permeability or plasma LPS levels in PL-induced mice, whereas hepatic LPS levels were noted to increase. Although we observed increased intestinal permeability of PL mice, the lack of statistical significance may be due to individual variations in gut permeability within the PL group. Based on our experimental design, we administered P-HFD mice with weekly intraperitoneal injections of LPS. The low-dose, weekly intraperitoneal injection of LPS in our study is unlikely to have substantially affected plasma and hepatic LPS levels, as the observed levels were relatively low compared to the injection dose. Additionally, LPS in the circulation was found at a level similar to that in the control group, these data suggested that the elevation of hepatic LPS levels in the liver was derived from the gut and not the extra injection of LPS. LPS is a risk factor for inducing hepatic inflammation and NASH. The upregulation of the LPS-TLR4 pathway leads to NF-κB activation and inflammatory cytokine production, which play key roles in NASH progression and development (41). PL-induced NASH mice exhibited a higher expression of TLR4 and NF-κB, which resulted in a more severe inflammatory condition than mice supplemented with GEO. In addition to the TLR4/NF-κB pathway, LPS is also involved in activating the NLRP3 inflammasome (64).
We also conducted a dosage translation of GEO from mice to humans and compared it with the dosage achievable from natural ginger (Supplementary Fig. 5). Our analysis indicates that the dosage used in the study can be obtained from natural ginger, suggesting the potential development of GEO as a functional food or food supplement. Nevertheless, further investigation through human studies is required to comprehensively evaluate its potential applications. Histological analysis of kidney, spleen, and ileum sections in the GEO-treated group did not reveal any significant findings, as shown in Supplementary Fig. 6. These results indicate the safety of GEO under our experimental conditions. Moreover, a subchronic toxicity study conducted on Wistar rats for a duration of 13 weeks reported no observed adverse effect level (NOAEL) of 500 mg/kg body weight (65). When extrapolated to mouse dosage (66), this corresponds to 1000 mg/kg body weight. Importantly, it should be noted that the dosages utilized in our study were relatively lower than the NOAEL reported in the literature, providing further evidence of the safety profile of GEO at the selected dosages.
Collectively, the results of our study demonstrated that GEO exhibits a potential hepatoprotective effect by preventing NASH progression in a PL-induced NASH mouse model. The mechanisms of action of GEO include reducing fat accumulation, oxidative stress, hepatic inflammation, blocking the NLRP3 inflammasome pathway, remodeling the gut microbiota, and modulating the LPS/TLR4/NF-κB pathway. GEO can, therefore, be used as a dietary supplement to prevent NASH.
{kind=link}