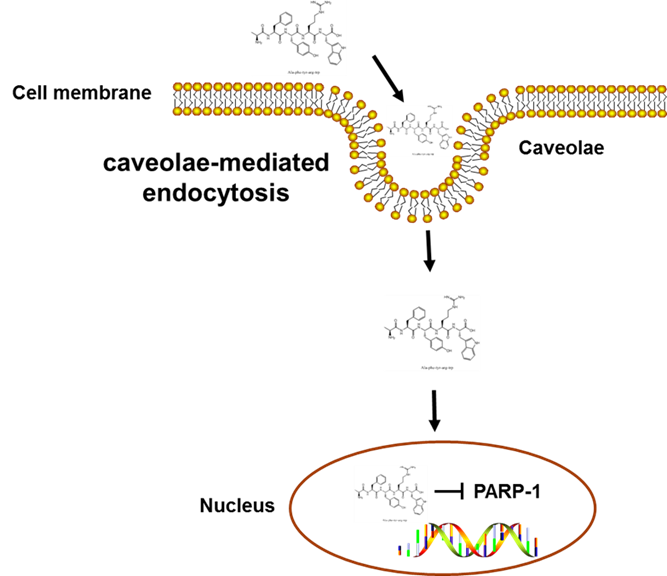
In this study, we employed the endocytosis pharmacological inhibition strategies and treated HepG2 cells with a variety of inhibitors to determine the specific mechanism by which Fagopyrum tataricum-derived peptide AFYRW enters cells. Endocytosis is an energy-dependent process in which extracellular substances (such as proteins, peptides, microorganisms, and nanoscale systems) are engulfed into the cell through deformation and movement of plasma membrane. Generally, endocytic pathways can be classified into two types: phagocytosis (for large nanoparticles) and pinocytosis (for fluids and solutes). Pinocytosis is usually divided into clathrin-dependent endocytosis (CDE) and clathrin-independent endocytosis (CIE). CIE involves three distinct mechanisms: the caveolae-mediated endocytic pathway, the clathrin/caveolae-independent endocytic pathway, and the macropinocytosis pathway. Clathrin/caveolae-independent endocytosis can be further divided into Cdc42, RhoA, flotillin, and Arf6 pathways [26]. Macropinocytosis, as a unique pathway of endocytosis, usually mediates the entry of macromolecules or viruses into cells. To prevent macropinocytosis, we exposed HepG2 cells to AMR, which suppresses macropinocytosis by reducing Na+/H+ exchange at the cell surface and blocking Rac1 and Cdc 42 signal transduction [27]. As expected, our results demonstrated that although cellular uptake of AFYRW required active endosomal acidification, AMR did not significantly alter FITC-AFYRW fluorescence intensity, indicating that macropinocytosis was not responsible for its cellular entry. The clathrin-dependent endocytosis pathway, which exists in all mammalian cells, is the primary way cells obtain nutrients. CPZ is a cationic amphipathic and the most important drug that inhibits the clathrin-dependent endocytosis pathway. CPZ relocates clathrin and adaptor protein complex-2 (AP2) from the cell surface to the intracellular endosomal membrane, causing the plasma membrane to collapse and form vesicles, thereby blocking CDE [28, 29]. When HepG2 cells were pretreated with CPZ, the uptake of AFYRW did not decrease, but instead increased, which may result from a compensatory phenomenon based on CDE changes [30]. It has been similarly reported in the literature that some individual treatments with endocytic inhibitors revealed higher cellular uptake compared to the untreated group [18]. It is possible that AFYRW enters cells via multiple pathways simultaneously, and disrupting one endocytic pathway might cause the upregulation of other active pathways. The effect of incubation time with endocytosis inhibitors may be cell type-dependent, and a careful optimization would thus be required for each in vitro system. Caveolin is rich in cholesterol and sphingolipid, so the endocytosis pathway mediated by caveolae can be studied by cholesterol-sensitive reagents such as methyl-β-cyclodextrin (m-β-CD) or nystatin. M-β-CD was commonly used as a caveolae-mediated endocytosis inhibitor by sequestering cholesterol, which in turn perturbed plasma membrane fluidity in lipid rafts. However, it has been reported with growing evidence that m-β-CD is not a specific inhibitor, as it also can also inhibit CDE and macropinocytosis [31]. We thus chose nystatin, which is more specific towards caveolae-mediated endocytosis. A significant and concentration-dependent reduction in the signal intensity of fluorescently positive cells was observed upon 12.5 or 50 µg/mL nystatin pretreatment, indicating a block in AFYRW uptake. Taken together, our data suggest that AFYRW enters HepG2 cells by relying on caveolae-mediated endocytosis. It would be interesting to further explore whether cells are also using an energy-dependent pathway that is unaffected by the chemical inhibitors tested in this study over the incubation period.
Bioactive macromolecular peptides entering the cell via endocytosis need to escape endosomes before they can be delivered to cytoplasm or organelles, and effective delivery of drugs at specific sites is a prerequisite for pharmacological action and therapeutic effect. Endocytosis is composed of multiple steps: an engulfed membrane embolus is first pinched off, and the endosomes are then delivered to specific vesicles that sort the contents for different destinations, which include diverse organelles, the extracellular environment, and other cells. Consistent with our observation of the subcellular localization of AFYRW, studies have shown that hydrophobic amino acids, such as Phe (F) or Trp (W) hydrophobic aromatic rings, are beneficial for endosome escape [32]. AFYRW contains the hydrophobic amino acids Ala (A), Phe, and Typ, as well as aromatic amino acids Tyr (Y) and Trp, which might contribute to its endosome escape and nuclear translocation, in addition to its antioxidant properties. Furthermore, basic amino acids are particularly prominent among the nuclear targeting sequences. Nuclear localization signals (NLPS), for instance, are a kind of short peptide rich in basic amino acids, whose net positive charge can interact with negatively charged DNA and promote the nuclear targeting of drugs [33]. Arg (R) in AFYRW with its positively charged guanidine may specifically form hydrogen bonds with the phosphate skeleton of DNA/RNA [34]. It would be interesting to explore the therapeutic potential of AFYRW to enhance nuclear delivery in the absence of cytotoxicity.
The peptide AFYRW may play a role in modulating nuclear function, and we utilized biotin-AFYRW-streptavidin affinity purification method to screen for nuclear proteins that interact with AFYRW. After protein spectrum analysis, PARP-1 was identified as the potential target of AFYRW. PARP-1 is a key ribozyme commonly expressed in eukaryotic cells and plays a vital role in DNA damage and repair. When the DNA damage is mild, PARP-1 participates in the process of DNA repair. In the context of high expression of PARP-1 in tumor cells, inhibition of PARP-1 blocks DNA repair, promotes tumor cell apoptosis, and exerts anti-tumor activity [35]. When DNA damage is severe, PARP-1 is over-activated, which leads to the consumption of NAD+ and ATP; energy deficiency will then slow down glycolysis and cause mitochondrial dysfunction and cell death. Oxidative stress is the main reason for the induction of apoptosis, tissue damage, and pathological changes in the body. Therefore, inhibiting the activity of PARP-1 has become promising for the treatment of neurodegenerative [36], cardiovascular [37], and metabolic diseases [38]. However, currently marketed PARP-1 inhibitors have adverse side effects, and highly potent and selective drugs remain to be discovered. AFYRW has a demonstrated capacity for eliminating radicals [12]; its effectiveness as food-derived peptidic antioxidant prepared from enzymatic hydrolysis of edible protein warrants further investigation given its safety characteristics as a phytocompound.
Peptide drugs can mimic protein-binding domains and competitively inhibit protein-protein interactions with advantages over traditional small molecular inhibitors. The characterization of peptide-protein interactions is necessary for the purposes of clarifying important biological processes and designing peptides or peptidomimetic drugs. Molecular docking is a powerful computing tool that is widely used to predict the binding structure of protein-peptide complexes and can provide leads for targeted inhibitor design. The current peptide-protein docking algorithms can be divided into local and global peptide docking [20, 39]. HPEPDOCK developed a hierarchical flexible global peptide docking method through rapid peptide conformation generation and global docking, which performs well in many protein-peptide complex docking schemes. Therefore, we used HPEPDOCK to determine whether PARP-1 interacts with AFYRW. Consistent with the biochemical results, the docking analyses revealed that AFYRW interacted with eight amino acid residues of PARP-1 mainly through hydrogen bonds and pi–pi accumulation. So far, most of the PARP-1 inhibitors in clinical and preclinical trials are structural analogues of NAD+, which are designed to compete with NAD+ at the enzyme active site and bind to the catalytic domain of the enzyme to inhibit self-modification [23]. Our data showed that AFYRW interacts with several key amino acids (Ala898, Tyr907, Lys903, and Asp766) that were reported to inhibit the activity of PARP-1 [25], confirming that AFYRW may be a potential inhibitor of PARP-1. The antioxidant mechanisms of food-derived peptides can be classified into three types: regulation of reactive oxygen species production, inhibition of enzymatic activity, or modification of the antioxidant pathway. Our study suggests the molecular regulation of PARP-1 enzyme activity; however, AFYRW may act on multiple pathways to exert its antioxidant activities.
There are several limitations to this study. First, we only used HepG2 cells as the target and alternative cell lines were beyond the scope of the current study. Second, although our data imply that AFYRW could be a potential PARP-1 inhibitor, the precise mechanisms of AFYRW function remains to be elucidated. Another challenge was that the bioactivity of AFYRW was tested via in vitro biochemical assays and cell culture, which should be verified by in vivo studies through animal models and human clinical trials. Hence, further work is needed to address these limitations.
{kind=link}