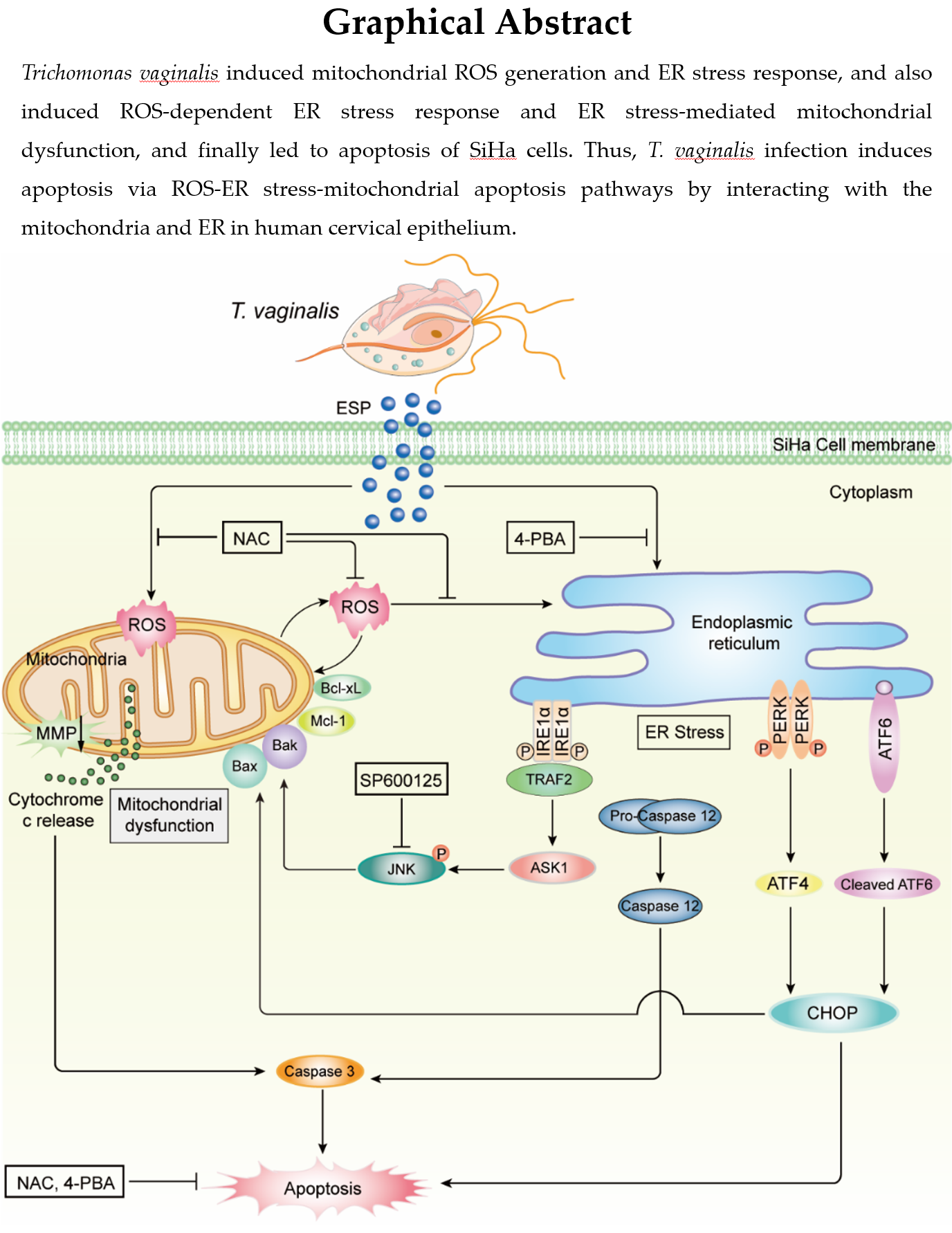
T. vaginalis induced cytotoxicity and apoptosis in SiHa cells in a parasite burden- and infection time-dependent manner
To assess the cytotoxicity of live T. vaginalis trophozoites in cervical epithelial SiHa cells, the cells were infected with T. vaginalis at different multiplicity of infection (MOI) and infection time before conducting the LDH assay. The cytotoxicity of SiHa cells infected with T. vaginalis at MOI 2 was 13.5 ± 1.1%, 26.9 ± 2.7%, and 46.9 ± 1.7% at 0, 2, and 6 h post-infection, respectively. Further, we observed that the toxicity of T. vaginalis in the SiHa cells was parasite burden-dependent, as it significantly increased to 48.9 ± 2.8% and 83.2 ± 5.0% at 2 and 6 h post-infection of T. vaginalis (MOI 5), respectively (Fig. 1A).
To determine whether this cytotoxicity was associated with apoptosis, we evaluated the apoptosis indicator proteins in the T. vaginalis-infected SiHa cells. The expression levels of cleaved PARP and caspase 3, as observed by western blotting, significantly increased from 2 h after T. vaginalis infection at MOI 2 in a parasite burden- and infection time-dependent manner (Fig. 1B).
Together, these results suggested that T. vaginalis induces cytotoxicity and apoptosis in SiHa cells in a parasite burden- and infection time-dependent manner.
T. vaginalis led to the generation of mitochondrial ROS in SiHa cells in parasite burden- and infection time-dependent manner
Oxidative stress is an important factor for cytotoxicity associated with apoptosis [17]. To investigate ROS production in T. vaginalis-infected SiHa cells, cellular and mitochondrial ROS levels were measured following infection. Cellular ROS levels significantly increased in the T. vaginalis-infected SiHa cells (MOI 2) from 2 h after infection and increased further at 6 h post-infection. Cellular ROS levels were significantly higher in the cells infected with T. vaginalis at MOI 5 than in cells infected with T. vaginalis at MOI 2 (Fig. 2A).
We further investigated mitochondrial ROS production using the MitoSOX Red staining technique and found that ROS production significantly increased from 2 h after T. vaginalis infection in direct proportion to parasite burden and infection time in the SiHa cells, which was similar to that in case of cellular ROS production (Fig. 2B).
These findings suggested that T. vaginalis induces mitochondrial ROS production in SiHa cells in a parasite burden- and infection time-dependent manner.
T. vaginalis induced mitochondrial apoptosis through ROS in SiHa cells
To determine whether ROS are involved in the induction of apoptosis in T. vaginalis-infected SiHa cells, the cells were pretreated with the ROS scavenger N-Acetyl cysteine (NAC) and the expression of apoptosis-related proteins and MMP was evaluated. We first determined the effects of NAC on cell viability using the MTS assay. Treatment of the SiHa cells with 0.2–1 mM NAC for 6 h elicited no significant differences in cell viability compared with those in the medium-treated group; however, the SiHa cells treated with 5 mM NAC for 6 h showed slightly reduced viability (see Additional file 1: Fig. S1A). Both cellular and mitochondrial ROS production were significantly suppressed in the T. vaginalis-infected SiHa cells following NAC pretreatment in a concentration-dependent manner (see Additional file 2: Fig. S2A, B).
As shown in Fig. 3A, in the NAC-pretreated SiHa cells, T. vaginalis-induced cleavage of PARP and caspase-3 was significantly suppressed in a dose-dependent manner. The permeabilization of the mitochondrial outer membrane is a crucial step in the progression of apoptosis [6, 7]; thus, we also investigated whether the T. vaginalis-induced ROS-dependent apoptosis in SiHa cells was related to the mitochondrial apoptotic pathways. JC-1 staining, an indicator of MMP. T. vaginalis infection, revealed a notable decrease in MMP levels, as indicated by the reduction in red fluorescence signal and an increase in green JC-1 fluorescence signal, in a parasite burden- and infection time-dependent manner compared with the signals in untreated control cells (Fig. 3B). However, these changes in JC-1 dye fluorescence were suppressed by NAC pretreatment in a dose-dependent manner (Fig. 3C).
These results suggested that T. vaginalis induces mitochondrial apoptosis through ROS, and ROS act upstream of PARP and caspase-3 during apoptosis induction.
T. vaginalis induced ER stress response and ER stress-mediated apoptosis in SiHa cells.
ER stress is known to contribute to apoptosis [18, 19, 21]. First, to check whether T. vaginalis causes ER stress in SiHa cells, we investigated the expression of ER stress-related proteins in T. vaginalis-infected SiHa cells. As shown in Fig. 4A, Ero1-Lα, p-IRE1α, CHOP, p-PERK, p-elF2α, ATF4, and cleaved ATF6 protein levels were significantly increased in the T. vaginalis-infected SiHa cells in a parasite burden- and infection time-dependent manner, whereas procaspase-12 levels were decreased as compared with those in the untreated control group (Fig. 4A).
Next, to confirm whether apoptosis is induced by ER stress in T. vaginalis-infected SiHa cells, the cells were pretreated with the ER stress inhibitor 4-phenylbutyric acid (4-PBA), and the expression of ER stress- and apoptosis-related proteins was evaluated. The SiHa cells treated with 5 mM 4-PBA for 6 h did not show a significant difference in viability compared with that of the untreated control group (see Additional file 1: Fig. S1B). The expression of ER stress-related proteins was reversed in the 4-PBA-pretreated T. vaginalis-infected SiHa cells in a dose-dependent manner as compared with that in the untreated T. vaginalis-infected cells (Fig. 4B). In addition, 4-PBA pretreatment attenuated the levels of cleaved PARP and caspase-3 in T. vaginalis-infected SiHa cells in a dose-dependent manner (Fig. 4C). These results demonstrated that T. vaginalis induces ER stress response as well as ER stress-mediated apoptosis in SiHa cells.
T. vaginalis induced ROS-dependent ER stress responses in SiHa cells.
ER and mitochondria are closely related, both structurally and functionally [18, 19, 22, 23]. Thus, to evaluate the effects of mitochondrial ROS in the induction of ER stress response in T. vaginalis-infected SiHa cells, the SiHa cells were pretreated with NAC, and the expression of ER stress-related proteins was investigated. Pretreatment with NAC significantly reduced the expression of Ero1-Lα, p-IRE1α, CHOP, p-PERK, p-elF2α, ATF4, and cleaved ATF6 in the T. vaginalis-infected SiHa cells in a dose-dependent manner, whereas the expression level of procaspase-12 was increased in a dose-dependent manner (Fig. 5A). Confocal microscopy imaging also confirmed that NAC pretreatment reduced T. vaginalis-induced upregulation of CHOP protein expression in a dose-dependent manner (Fig. 5B).
These results indicated that T. vaginalis induces ER stress response through mitochondrial ROS in SiHa cells, which leads to apoptosis.
T. vaginalis induced ER stress-mediated mitochondrial dysfunction in SiHa cells.
We also investigated whether ER stress affects mitochondrial functions in T. vaginalis-infected SiHa cells. Members of the Bcl-2 family are major regulators of mitochondrial integrity and mitochondria-dependent caspase activation, and MMP is the key indicator of mitochondrial apoptosis [6, 7]. Therefore, we investigated mitochondrial functions, including alternation of MMP and Bcl-2 family members, in T. vaginalis-infected SiHa cells. JC-1 dye staining revealed that T. vaginalis induced MMP depolarization in the SiHa cells; however, these changes in fluorescence were suppressed by 4-PBA pretreatment (Fig. 6A).
The expression of p-Bad (Ser112), Bax, Bak, Bik, and Puma was significantly increased in the T. vaginalis-infected SiHa cells (Fig. 6B), whereas the Bcl-xL and Mcl-1 levels were decreased in a parasite burden-dependent manner (Fig. 6C). The expression levels of p-Bad (Ser 112), Bax, Bak, Bik, and Puma were significantly decreased in the 4-PBA-pretreated T. vaginalis-infected SiHa cells in a dose-dependent manner (Fig. 6D), whereas the levels of Bcl-xL and Mcl-1 proteins were increased (Fig. 6E). These results clearly indicated that T. vaginalis induces mitochondrial dysfunction via ER stress in SiHa cells, leading to apoptosis.
T. vaginalis induced ER stress-mediated mitochondrial apoptosis via the IRE1/ASK1/JNK/Bcl-2 family protein pathways in SiHa cells.
Previous studies have reported that the IRE1/ASK1/JNK signaling cascade is one of the main downstream targets for the regulation of ER stress-induced cell apoptosis [18, 19, 21–23]. Therefore, we investigated whether T. vaginalis activates ER stress-dependent apoptosis through the ASK1/JNK pathway in SiHa cells. We found a substantial increase in the phosphorylation levels of both ASK1 (Ser83, Ser967, and Thr845) and JNK in the T. vaginalis-infected SiHa cells in a parasite burden- and infection time-dependent manner (Fig. 7A). Next, we investigated whether ASK1/JNK activation was related to ER stress in the T. vaginalis-infected SiHa cells. Western blotting analysis revealed that 4-PBA pretreatment attenuated the T. vaginalis-induced elevation of ASK1 and JNK phosphorylation levels in a dose-dependent manner (Fig. 7B). These data indicated that the T. vaginalis-induced ER stress activates the ASK1/JNK pathway in SiHa cells.
To evaluate whether the ASK1/JNK pathway is associated with mitochondrial apoptosis in T. vaginalis-infected SiHa cells, the cells were preincubated with the JNK inhibitor SP600125, and expression of MMP, Bcl-2 family proteins, and apoptosis-related proteins was investigated. The SiHa cells treated with 30 µM SP600125 for 6 h did not display a significant difference in their viability as compared with those of the untreated control group (see Additional file 1: Fig. S1C). Western blotting analysis revealed that Bak, Bax, cleaved PARP, and caspase-3 levels decreased in a dose-dependent manner in the SP600125-pretreated T. vaginalis-infected SiHa cells (Fig. 7C). In contrast, Bcl-xL and Mcl-1 levels were increased by pretreatment with SP600125 in the T. vaginalis-infected SiHa cells in a dose-dependent manner (Fig. 7D). In addition, SP600125 pretreatment substantially increased MMP levels in the T. vaginalis-infected SiHa cells, as demonstrated by increased red and reduced green JC-1 fluorescence, in a dose-dependent manner (Fig. 7E). These observations provided evidence that T. vaginalis induces ER stress-mediated mitochondrial apoptosis via the IRE1/ASK1/JNK/Bcl-2 family protein signaling pathways in SiHa cells.
{kind=link}