Radiation combined with PD-1 blockade has significant anti-tumor effects.
To verify the roles of radiation combined with PD-1 blockade on progress of NSCLC, mice bearing xenografts were implanted by subcutaneously injecting LLC derived CSCs. We used flow cytometry to isolate CSCs from LLC by related markers- CD44 and CD133 (Fig. 1A). The tumors were monitored and were isolated. As shown in Fig. 1B and 1C, the tumor sizes were smallest in Radio + anti-PD-1. PET-CT results showed that compared with the other three groups, the Radio + anti-PD-1 had the lowest tumor metabolic value (Fig. 1D). H&E staining showed that the necrotic area of tumor in Radio + anti-PD-1 was much smaller, even if there were some decreases in Radio group (Fig. 1E). Consistent with H&E results, the apoptosis in Radio + anti-PD-1 was the highest compared with the other three groups (Fig. 1F). These results indicated that radiation combined with PD-1 blockade inhibited NSCLC effectively. To investigate the effect of radiotherapy with or without PD-1 inhibitor on immune microenvironment of NSCLC, we examined CD3 and CD8 positive cells in tumor tissue. The results showed that the number of CD3 + and CD8 + cells increased significantly in Radio + PD-1 (Fig. 1G). Then, peripheral blood T cells in each group were sorted, IFN-γ, CD44, IL-4, and CD28 in each group were detected by qRT-PCR to evaluate the activation status. The result showed that treated with radiotherapy or PD-1 blockade or both promoted IFN-γ, CD44, and CD28 levels (P < 0.05), while radiation combined with PD-1 blockade displayed the most significant increase (Fig. 1H). Results As shown in Fig. 1I, the number of CD4 and CD8 positive T cell, Treg and NK cell were increased after radiotherapy or PD-1 treatment alone or both.
Radiation combined with PD-1 alters the cell types and phenotype of CSCs in NSCLC by single-cell transcriptomics.
To explore intratumor heterogeneity in NSCLC, we used nanopores to capture single cells through the BD Rhapsody platform to generate matched single-cell transcriptome profiles of Model and Radio + PD-1 from two animals (Fig. 2A). After initial quality control, single-cell transcriptomes were available for a total of 21019 cells for further analysis. By analyzing the variably-expressed genes in all cells, we identified the eleven major cell types in the nineteen cell clusters by UMAP. These cells included fibroblast (cluster 1, 8, 13, 16, and 18), macrophages\monocyte (cluster 3, 5, and 17), B cell (cluster 9,19), DC (cluster 11), endothelial cells (cluster 12), neutrophils (cluster 15), T\NK cell (cluster 7), NK (cluster 14), and cancer stem cells (cluster 2, 4, 6, and 10). We then performed differential gene expression analysis to determine the identity of these cell clusters. Each cluster was compared with other aggregated clusters to find unique gene signatures, and the top 10 significantly differentially expressed genes for each cluster were shown in the heat map, as shown in Fig. 2B. The eleven cell types in Model and Radio + PD-1 were displayed by UMAP plot and Violin plots (Fig. 2C-E). Therefore, these results reveal that NSCLC is highly heterogeneous and composed of diverse cell types. To investigate the effects of radiation combined with PD-1 blockade on CSCs, we performed a cluster analysis of stem cells. Compared with the Model, radiation combined with PD-1 blockade significantly reduced the number of CSCs (P < 0.05) (Fig. 2F). IF showed that compared with the Model group, the enrichment of CD133 + cells (CSCs) were slightly lower in anti-PD-1 or Radio group and this enrichment was lowest in Radio + anti-PD-1 group (Fig. 2G). In addition, we also examined the CSC phenotype. RT-qPCR and Western blot were performed the detect the levels of OCT4, SOX2, and β-catenin in lung cancer tissues after radiation combined with or without PD-1 blockade. The results showed that the inhibition of OCT4 and SOX2 was most significant in Radio + anti-PD-1 group (P < 0.05) (Fig. 2H-J).
Radiation combined with PD-1 affects T-cell subgroup.
To depict their intrinsic portrait, these T cells (cluster 7) were further divided into six subclusters (Fig. 3A). Unique gene signatures and the top 10 SDE genes were outlined for each T-cell subgroup (Table S3 and Fig. 3B). Cell clusters were specifically classified according to different molecular features of T cells among tissue-stratified clonotypes (Fig. 3C). The UMAP plot displays seven subclusters of T cells (Fig. 3D) and compared in Model and Radio + PD-1(Fig. 3E). The ratio of different subclusters of T types (Fig. 3F) and statistical analysis (Fig. 3G) were carried out in Model and Radio + PD-1. The results showed that the proportion of each T cell subtype was significantly changed in the Radio + PD-1 group.
Differentiation status of CD8 + T cell and its role in cell communication.
The tumor cells were stratified during cancer development using pseudo-time reconstruction (Fig. 4A). Trajectories that predicted the path of de novo tumor progression showed T-cell distributions in two animals. The population of exhausted CD8 + T cells in Radio + PD-1 group decreased while the population of naive CD8 + T cells increased along the trajectory, validating the reliability of the trajectory analysis (Fig. 4B). Our data showed dynamic gene expression profiles during the malignant evolution of tumor cells (Fig. 4C). To investigate the relationship between different CD8 + T cell subtypes and their relationship with CSCs, we analyzed the intercellular communication network. The network diagram of the strength of cell-to-cell interaction showed that different subtypes of CD8 + T cells could convert to each other and were all affected by CSCs (Fig. 4D, E).
Radiation combined with PD-1 blockade increases m6A methylation level and targets on Wnt10b in lung cancer tissues.
As RNA m6A methylation modification plays critical roles in pathological and physiological processes, we hypothesized that RNA m6A methylation modification also existed in the therapy of lung cancer. To verify this, tumor tissues of the Model and Radio + PD-1 were collected for m6A sequencing. The results showed that m6A peaks increased in Radio + PD-1 compared with the Model group (Fig. 5A). Next, we extracted the differentially expressed genes from m6A-seq and RNA-seq and classified the genes according to hyper methylation and down transcription (Hyper-down), hyper methylation and up transcription (Hyper-up), hypo methylation and down transcription (Hypo-down), hypo methylation and up transcription (Hypo-up). The proportion of Hyper-down was even higher than the other three groups, which were displayed in a four-quadrant diagram (Fig. 5B). By analysis of the frequency of m6A-peak in CDS region and 3’UTR region in the Model and Radio + PD-1, it was found that the frequency of m6A-peak in 3’UTR region in the RT + PD-1 group was significantly increased (Fig. 5C). Next, the transcriptome sequencing data were further extracted, and the differentially expressed genes between the two groups were extracted with P-vale < 0.05 and log2 (fold change) > 0.58. The data were displayed by volcano diagram (Fig. 5D). There were significantly up-regulated and down-regulated genes. Followed by m6A - seq data extraction, screening P - Vale < 0.05 and | log2 (a fold change) | > 0.58 the differences between the two groups of m6A modified genes for data extraction, and through the volcano's figure display, the result is shown in Fig. 5E.
To determine the major gene in the m6A methylation, we extracted the differentially expressed genes with highly m6A methylation and transcription down. A total of 34 differential genes with increased m6A modification and decreased transcription levels were selected (Amica1, Angptl7, Ankrd35, Arhgap27, Batf2, Bend6, Camk2b, Chga, Doc2a, Efnb1, En2, Evl, Fat4, Flrt2, Gfi1, Gjb3, Gm40447, Jph2, Jsrp1, Kif26b, Lysmd1, Mmp28, Mmrn2, Nlrc5, Nrg2, Palmd, Plxna2, Ptger4, Rgs1, Runx3, Sectm1a, Tnnt 1, Wnt10b, Xirp1), and the m6A data of the above differential genes were extracted, and it was found that the m6A of Wnt10b was significantly increased (Fig. 5F, G). Given that methylation level increased in lung cancer tissues of Radio + anti-PD-1 group, we collected tumor tissues to explore the methylation level of Wnt10b by MeRIP-RT-qPCR. The results showed that compared with Model, the methylation levels of Wnt10b in the Radio group, PD-1 group, and Radio + anti-PD-1 group were all increased (P < 0.05), and the Radio + anti-PD-1 group had the highest methylation level of Wnt10b (Fig. 5H).
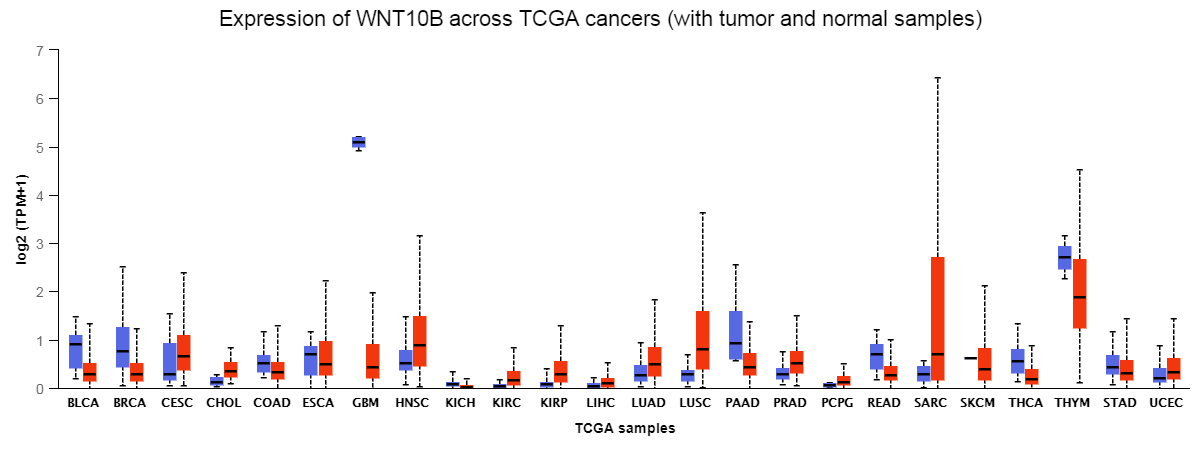
Additionally, ingenuity Pathway Analysis (IPA)-performed GO analysis of transcripts showed Wnt10b The top five pathways (by P value) are shown in Fig. 5I. Wnt10b was predicted to be highly expressed in tumors, especially in lung cancer, in the TCGA database (Supplementary Fig. S1). Western blot and IF verified that after treating with radiation combined with PD-1 blockade, the protein expression of Wnt10b decreased most compared with Model, anti-PD-1 or Radio group (Fig. 5J, K, O). To further investigate the role of Wnt10b in CSCs, we knocked down Wnt10B expression. The results showed that the inhibition of OCT4 and SOX2 was significant in sh-Wnt10b group (P < 0.05). Meanwhile, β-catenin showed the same trend as OCT4 and SOX2 in vitro experiments (Fig. 5L-N). Next, we explored tumor cell apoptosis in vivo. TUNEL assay result showed that cell apoptosis increased after Wnt10b interference, and the apoptosis was more obvious after combined with PD-1 inhibition (Fig. 5P). These results showed that Wnt10b is a major gene involved in CSC proliferation and apoptosis.
FTO is involved in CSC proliferation and apoptosis.
To find the key regulatory molecules responsible for radiation combined with PD-1 blockade, we performed RNA-seq to detect the differential expression genes in Model and Radio + anti-PD-1 groups. The results showed that the gene expression of FTO was significantly inhibited in Radio + anti-PD-1 group (Fig. 6A). Western blot and RT-qPCR were used to detect the expression of Methylases (METTL3, METTL14, WTAP), demethylases (FTO, Alkbh5), reader recognition proteins (YTHDF1, YTHDF2, YTHDF3, YTHDC1, YTHDC2) and Wnt5b. The results showed that the expression of methylases (METTL3, METTL14, WTAP) was increased (P < 0.05), while the expression of demethylases (FTO, Alkbh5) was decreased (P < 0.05) in the tumor-bearing mice after radiation combined with PD-1 blockade (Fig. 6B-D). The results showed that the methylation level of cancer mice might increase after radiation combined with PD-1 blockade, and the difference of the expression of the marker demethylase FTO was the most obvious.
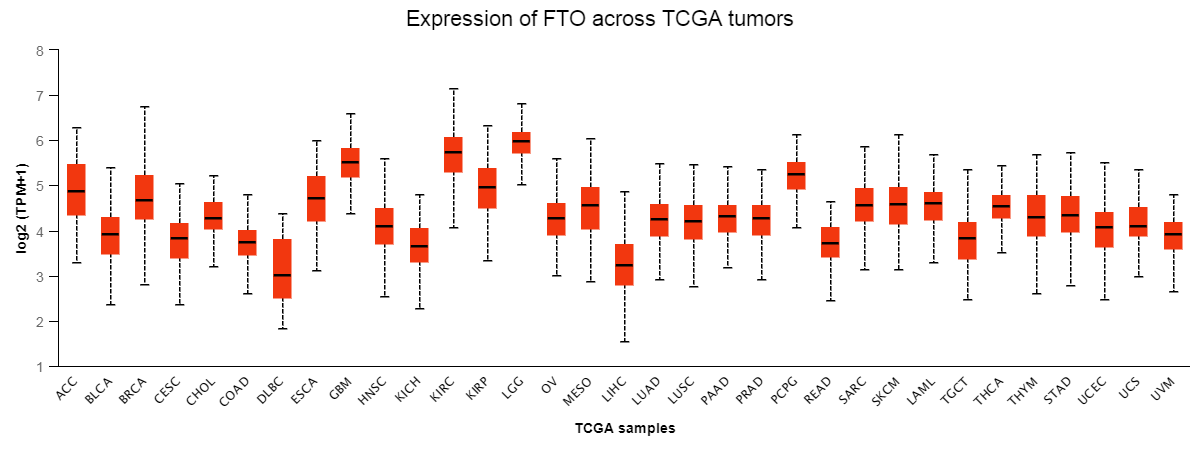
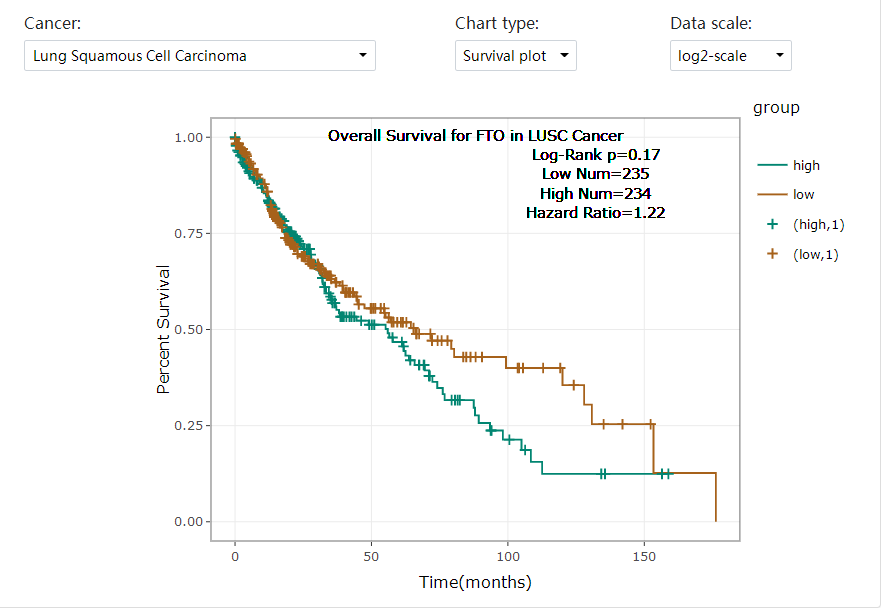
In order to verify the existence of mRNA base modification behavior during treatment, we used colorimetric method to test the m6A level in the tissues of each group and compared the m6A modification status among the groups. Results As shown in Fig. 6E, compared with the Model group, tissue methylation occurred in both the Radio group and PD-1 group, and the overall degree was not high, and the methylation level was significantly increased after combined treatment (P < 0.05). In TCGA tumor dataset, expression of FTO in lung cancer was highly expressed (Supplementary Fig. S2). Analysis combined with Kaplan-Meier survival analysis and TCGA dataset were performed found that five-year survival rate was much higher in patients with low level of FTO compared with those with high level of FTO (Supplementary Fig. S3). These findings indicated that FTO expression was associated with patient survival and might contribute to cancer progression. To evaluate the expression of FTO in different cell line of lung tissue, we first detected the relative expression level of FTO in lung cancer cells (LLC, LLC-CSCs, A549 and A549-CSCs) and human lung bronchial epithelial cells (BEAS-2B). Compared with BEAS-2B cells, FTO expression was higher in lung cancer cells (LLC, LLC-CSCs, A549 and A549-CSCs) and those in LLC-CSCs was the highest (Fig. 6F). Thus, LLC-CSCs might be the most effective target for FTO which was the main target in this study.
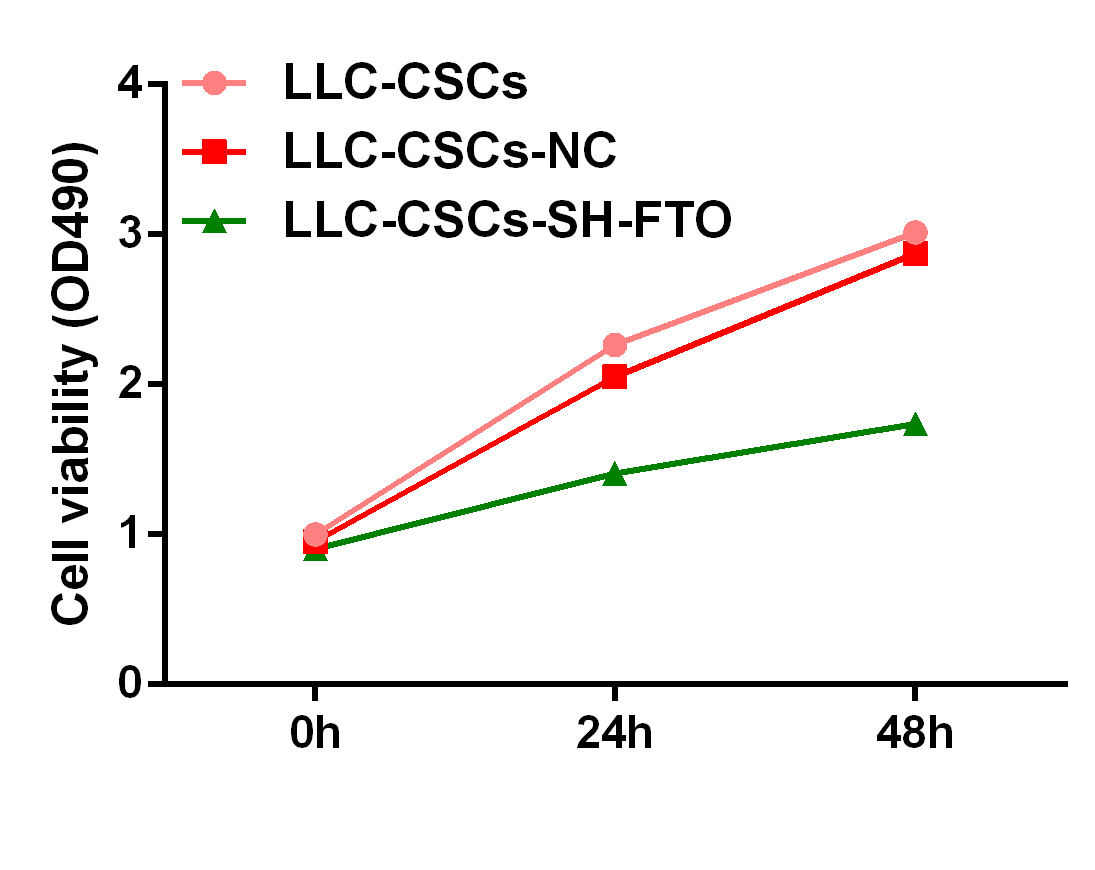
As FTO was highly expressed in CSCs and associated with poorly patient survival, we investigated the roles of FTO in NSCLC. We synthetized shRNA to knock down FTO which was knocking down FTO expression effectively in LLC-CSCs (Fig. 6G, H). MTT showed that knocking down FTO expression inhibited cell viability of LLC-CSCs (Supplementary Fig. S4). Colony formation assay showed that compared with the control group, sh-FTO inhibited colony formation of LLC-CSCs (Fig. 6I). we also found that knocking down of FTO promoted cells apoptosis and cell cycle arrest through Tunel and flow cytometry (Fig. 6J-L). In addition, we found that sh-FTO inhibited sphere formation of LLC-CSCs (Fig. 7M). These results indicated that FTO is involved in CSC proliferation and apoptosis.
To further verify the role of FTO on tumorigenesis, CSCs transfected with FTO virus were inoculated to establish an animal model. The tumor formation and tumor size of the animals are shown in Fig. 6N and 6O. The tumor sizes were smaller in Radio + anti-PD-1 and siFTO. The TUNEL assay showed that apoptosis was the highest in Radio + anti-PD-1 and siFTO compared with the other two groups (Fig. 6P). In addition, IF results showed that CD8 + T cells increased significantly in Radio + PD-1 (Fig. 6Q). While CD133 + cell significantly increased in OE-FTO group and significantly decreased in Radio + anti-PD-1 group (Fig. 6Q). To further evaluate the effect of FTO on T cells activation, we performed flow cytometry to analyzed CD4+, CD8+, Treg, and NK cells after FTO interference or overexpression. The results showed that CD8 + T cells decreased significantly in OE-FTO group, whereas cells increased significantly in siFTO group and Radio + anti-PD-1 group (Fig. 6R). These findings demonstrated that radiation combined with PD-1 blockade has significant anti-tumor effects by downregulating FTO expression. Additionally, we explored how FTO regulates CSC stemness. RT-qPCR and Western blot results showed that the inhibition of OCT4 and SOX2 in siFTO group and Radio + anti-PD-1 group (P < 0.05) (Fig. 9K-M). Meanwhile, β-catenin showed the same trend as OCT4 and SOX2 in vivo and in vitro experiments (Fig. 6S-U). These findings demonstrated that radiation combined with or without PD-1 blockade regulated CSC stemness through downregulating FTO.
Wnt10b promotes FTO RNA methylation after treating with radiation combined with PD-1 blockade.
In addition, we found that the methylation level of Wnt10b was significantly increased in the CSCs-SH-FTO group (Fig. 7A). Further, knocking down of FTO led to a decrease both in pre-mRNA- Wnt10b and mature mRNA- Wnt10b levels (Fig. 7B, C). In addition, we verified this by examining tumor tissue in animals. FTO interference vector was constructed, and the expression levels of FTO and Wnt10b in tumor tissues were detected by WB and qPCR. The results showed that the transcription and translation levels of Wnt10b was decreased after interfering with FTO, further confirming that FTO inhibited Wnt10b expression (P < 0.05) (Fig. 7D. E). These results showed that the expression of FTO and Wnt10b was positively correlated. These results indicated that FTO could binding Wnt10b m6A to inhibit its translation.
{kind=link}
{kind=link}
{kind=link}
{kind=link}