KPT-9274 is a potent and selective NAMPT inhibitor.
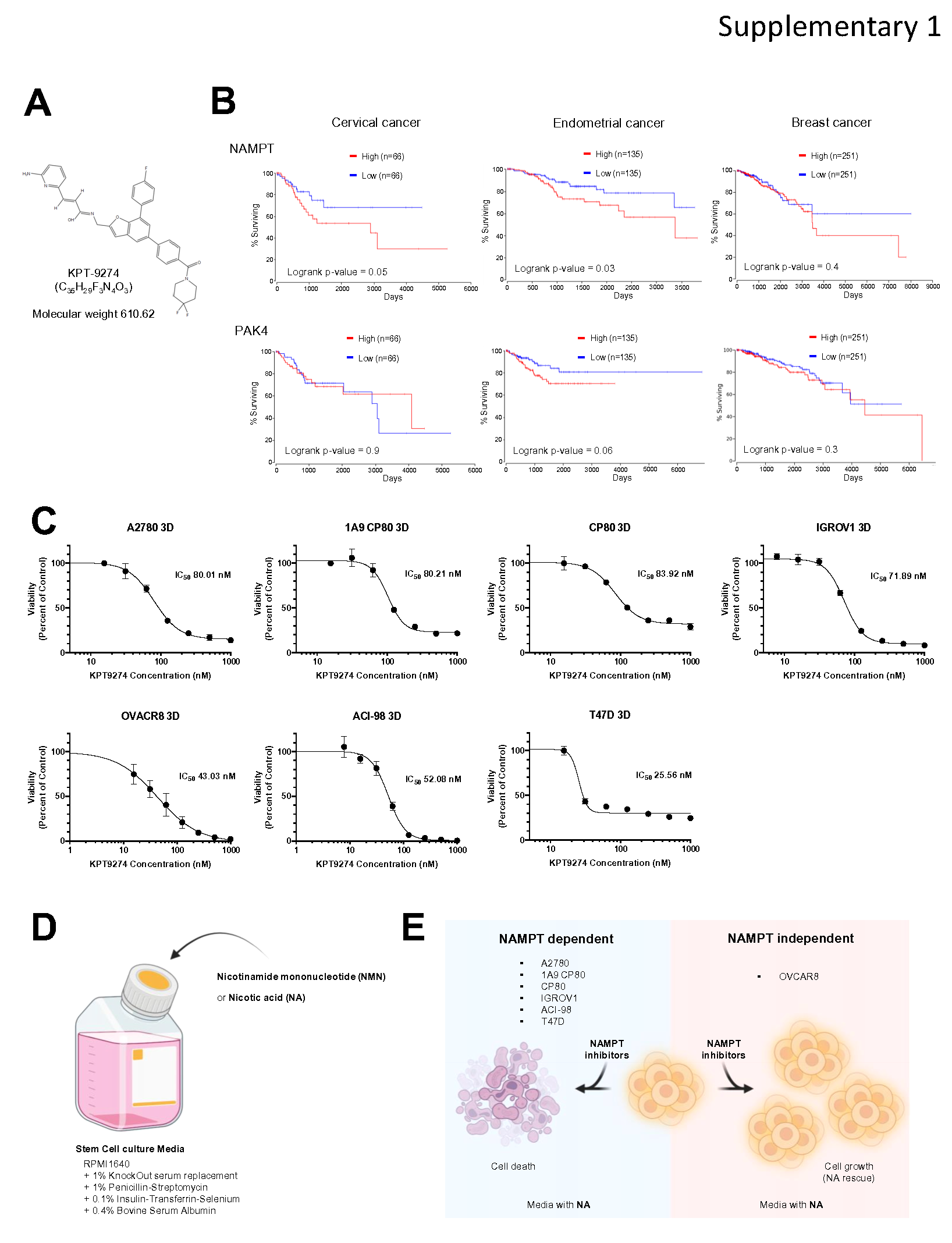
To investigate the relevance of NAMPT and PAK4 in ovarian cancers, we first examined TCGA datasets. High expression of NAMPT was correlated with a significant reduction in overall survival in human ovarian cancer, suggesting that high NAMPT expression may be a prognostic factor in ovarian cancer. Similar results were observed in cervical and endometrial cancers, but there was no significant difference in breast cancer. High PAK4 expression lacked strong negative prognosis in these cancers, although a trend towards worse outcomes existed in endometrial cancer. (Fig. 1C and Supplementary Fig. 1B).
To assess preclinical effectiveness of KPT-9274 in gynecological cancers, we tested the effect on cell viability of 3D spheroids from 11 cell lines of different histologic subtypes. The cell lines we used in this study had varying degrees of sensitivity to KPT-9274, and differed in their ability to be rescued by NMN or NA addition (Table 1). Based on manufacturer's recommended concentrations and previous reports, KPT-9274 was tested up to 1,000 nM as the highest concentration [24] [25] [26] [27] [31]. The efficacy of KPT-9274 was demonstrated against A2780, 1A9CP80, CP80, IGROV1 and OVCAR8 in ovarian cancer, ACI-98 in endometrial cancer and T47D in breast cancer with IC50 25–83 nM. In contrast, KPT-9274 did not inhibit the viability of SKOV3, EFE-184, KLE and MCF-7 at the highest dose, indicating that these cells are not primarily dependent on NAMPT in NAD+ synthesis (Supplementary Fig. 1C). As expected, addition of NMN (downstream of NAMPT) rescued KPT-9274 impact across all cell lines (NMN rescue). To further test whether the cell lines produced NAD+ from NA by other pathways, rescue experiments were performed. We observed that NA, but not NMN, rescued the cytotoxic effect of KPT-9274 in OVCAR8 (NA rescue) (Fig. 1D and Supplementary Fig. 1D). Notably, NAD+ production pathway differed across the different cell lines. While multiple ovarian cancer cell lines are dependent on NAMPT for NAD+ production, therefore respond to KPT-9274, OVCAR8 does not rely on NAMPT for NAD+ synthesis and uses alternative pathways to produce NAD+. Meanwhile, SKOV3, EFE-184, KLE were resistant to KPT-9274 (Table 1). This suggests that classification of the ovarian cancer patients based on NAD+ biosynthesis pathway is required for clinical application of KPT-9274 (Supplementary Fig. 1E).
Next, to examine KPT-9274's potential in platinum-resistant ovarian cancers, we tested using different cell lines, including platinum-sensitive (A2780) and platinum-resistant sub-lines (1A9CP80 and CP80). Based on clinical studies that reported the blood concentration of cisplatin [32] [33], the maximum concentration of cisplatin in this experiment was set at 20 µM. KPT-9274 demonstrated similar anti-tumor effects to cisplatin on A2780 (Fig. 1E). Notably, we observed KPT-9274 was significantly more effective than cisplatin in 1A9CP80 and CP80. Therefore, KPT-9274 could be a promising treatment for ovarian cancer that has developed resistance to platinum-based therapies.
KPT-9274 suppresses the production of NAD + , NADPH, and ATP.
To assess KPT-9274 impact on NAMPT-dependent cell lines, we first measured NAD+ and NADPH production at various concentrations. Using 3D-cultured CP80, ACI-98, and IGROV1, KPT-9274 inhibited NAD+ and NADPH production in a dose-dependent manner (Fig. 2A and 2B).
To further investigate the mechanism, we next tested KPT-9274 on ATP production, as NAD+ is essential for ATP generation through glycolysis and the TCA cycle [34]. Consistent with NAD+ and NADPH production, KPT-9274 treatment significantly reduced ATP levels (Fig. 2C). Together, we show that KPT-9274 inhibits NAD+, NADPH, and ATP production in multiple NAMPT-dependent ovarian cell lines, indicating that KPT-9274 induces a comprehensive disruption of vital biological processes.
KPT-9274 causes the suppression of mitochondrial function.
Based on the inhibitory effect on NAD+, NADPH and ATP, we next hypothesized that KPT-9274 affects mitochondrial functions. Using the Mito Stress Test with XFe96, we assessed KPT-9274 impact on mitochondria function in 3D-cultured CP80 and ACI-98. As anticipated, KPT-9274 reduced oxygen consumption rate (OCR), an established measure of mitochondrial function [35], in CP80 and ACI-98 cells in 3D spheroids, affecting both basal and maximal respiration (Fig. 3A and 3B). Interestingly, KPT-9274 significantly suppressed not only OCR, but also maximal extracellular acidification rate (ECAR), reflecting glycolysis (Fig. 3C). NAD+ is a co-enzyme in the reaction catalyzed by Glyceraldehyde 3-phosphate dehydrogenase (GAPDH), which is an enzyme essential for the conversion of glyceraldehyde-3-phosphate to 1,3-bisphosphoglyceric acid in glycolysis [36]. Hence, we hypothesized KPT-9274 inhibits GAPDH. As anticipated, KPT-9274 inhibited the GAPDH-mediated reaction, and adding NMN to the medium reversed the inhibition (Fig. 3D). These findings suggest KPT-9274 suppresses not only mitochondrial ATP production, but also glycolysis.
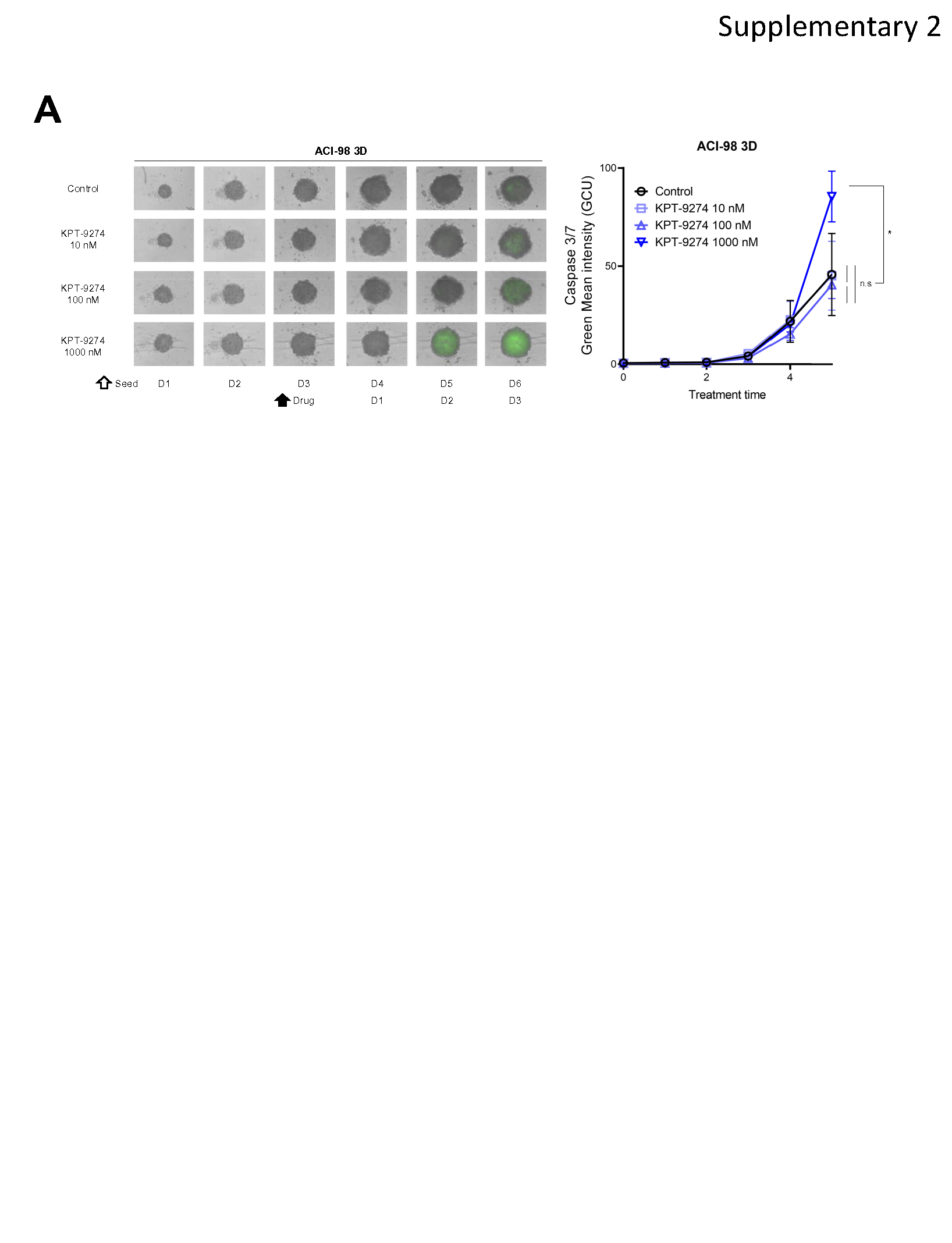
Furthermore, we investigated the impact of KPT-9274 on mitochondrial membrane potential, using TMRM, a fluorescent dye that is readily sequestered by active mitochondria. KPT-9274 significantly suppressed TMRM in CP80 and ACI-98 cells in 3D spheroids (Fig. 3E). Conversely, we observed up-regulated MitoSOX™ Red which reflects reactive oxygen species (ROS) generated in mitochondria of live cells (Fig. 3F). Moreover, cleaved caspase-3/7 signal was monitored over time using IncuCyte™ Caspase-3/7 Green Dye. KPT-9274 treatment significantly up-regulated green fluorescence intensity per area of spheroid, indicating caspase 3/7 activity was induced by KPT-9274 (Fig. 3G and Supplementary Fig. 2A). To evaluate whether KPT-9274 induces cell death, we quantified viability, cytotoxicity, and apoptosis induction using ApoTox-Glo™ Triplex Assay Kit, with or without a pan-caspase inhibitor, benzyloxycarbonyl-Val-Ala-Asp-fluoromethyl ketone (Z-VAD-FMK) [37], after 96 h of KPT-9274 treatment at varying doses. As anticipated, KPT-9274 suppressed cell viability and induced cytotoxicity as well as cleaved caspase-3/7 activity (Fig. 3H and 3J). Pre-treatment with Z-VAD-FMK inhibited only cleaved caspase-3/7 secretion, while having no considerable effect on cytotoxicity (Fig. 3I and 3J). These results suggest that caspase-3/7 activity is a part of anti-tumor effects of KPT-9274, but not entirely attributed to the cytotoxicity.
NAMPT correlates with inflammatory gene expression and PAK4 is associated with DNA repair genes in ovarian cancer patients.
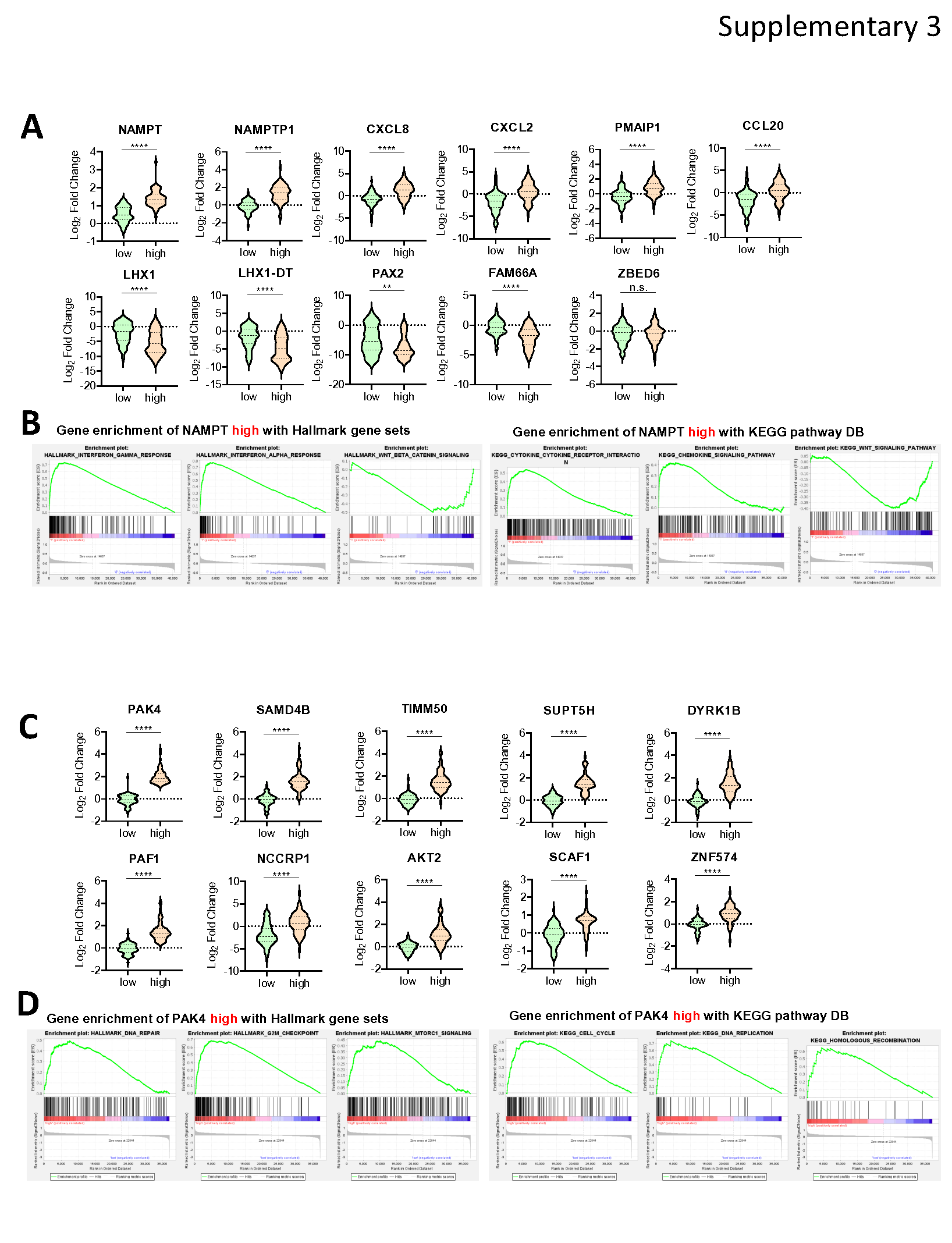
To further characterize the impact of NAMPT and PAK4 in ovarian cancer, we evaluated the ovarian cancer RNA sequencing data from TCGA. We first compared patients with high and low NAMPT expression (Lower percentile = 25% [n = 73), Upper percentile = 25% [n = 73)) and developed a heatmap and volcano plot to detect DEGs (Fig. 4A and 4B). Top 20 DEGs between NAMPT high and low expression patients included NAMPT, NAMPTP1, ARMC10, CAPZA2, CXCL8, CCDC71L, NCOA7, PMAIP1, SYPL1, PNPLA8, CXCL2, CEBPD, CCL20, ZBED6, FAM66A, PNMA8B, PYCR2, PSMC2, SOD2, and STEAP1 (Supplementary Fig. 3A and Supplementary Table 1). GSEA revealed that patients with high NAMPT expression exhibited enriched gene sets related to inflammation in Hallmark gene sets and KEGG pathway database. Top five up-regulated gene sets in NAMPT-high patients were TNF-α signaling via NFκB, Interferon-γ response, Interferon-α response, and Apoptosis in Hallmark gene sets. Moreover, using KEGG pathway, top five up-regulated gene sets were Cytokine-cytokine-receptor interaction, Chemokine signaling pathway, JAK-STAT signaling pathway, Nicotinate and nicotinamide metabolism, and Apoptosis (Fig. 4C and Supplementary Fig. 3B). Notably, the findings highlighted a connection between high NAMPT expression and increased inflammation, suggesting this may contribute to a poorer prognosis in ovarian cancer patients.
Similarly, we next identified DEGs in patients with high and low PAK4 expression (n = 73 and top 25%, respectively). A heatmap and volcano plot visualized results, highlighting 20 significant DEGs between PAK4 high and low expression patients including PAK4, POLR2I, ECH1, CAPNS1, PTOV1, RBPJ, ZNF628, YIF1B, KPNA5, RPL26P6, NDUFS7, ZNF865, ZC3H3, FRA10AC1, ZNF574, LY96, FRG1, MRPL2, C19orf47, and PPDPF (Supplementary Fig. 3C and Supplementary Table 2). All 23 DEGs were highly expressed genes in high PAK4 patients (p < 1e-11) (Fig. 4D and 4E). GSEA revealed that the top five up-regulated gene sets were G2M checkpoint, DNA repair, MTORC1 signaling, Wnt/β-Catenin signaling, and PI3K-AKT-MTOR signaling in Hallmark gene sets. Moreover, top five up-regulated gene sets were Cell cycle, DNA replication, Mismatch repair, Base excision repair, and Homologous recombination in KEGG pathway (Fig. 4F and Supplementary Fig. 3D). Collectively, these findings suggest elevated gene repair and cell proliferation functions in high PAK4 patients, potentially contributing to tumor cell survival and replication.
KPT-9274 triggers suppression of inflammatory signaling.
We hypothesized that the anti-tumor effects of KPT-9274 arise from inhibition of gene expression related to inflammation, gene repair, and cell proliferation signaling. To validate this hypothesis, we performed RNA-seq analysis on 3D cultured CP80 cells treated with DMSO (Control) or KPT-9274 for 24 h. First, principal component analysis (PCA) demonstrated that technical replicates in each group clustered together, indicating low variation between the replicate samples (Fig. 5A). Next, we conducted hierarchical clustering analysis to detect the DEGs based on RNA-seq data and constructed a heatmap and volcano plot to visualize the impact of KPT-9274 treatment. The top 20 DEGs between Control and KPT-9274 treatment were CA14, NLGN3, SCARA5, HDGF, NQO1, HMGA2, ERP27, HSD17B7, PPP2R5B, MYOF, PYM1, CDC42EP4, ACTA2, NQO2, YIPF6, ATXN2, PTMA, SLC30A8, SCN9A, and ZBTB2 (Supplementary Table 3). Interestingly, SNHG25, known for promoting ovarian cancer progression [38], and TMEM52B, associated with EGFR and E-cadherin modulation and tumor/metastasis suppression [39], significantly decreased with KPT-9274 treatment (Fig. 5B and 5C). Next, GSEA revealed top five up-regulated gene sets in Control compared to KPT-9274 treatment: Myc-targets-V1, Hedgehog signaling, Epithelial mesenchymal transition, Allograft rejection, and Interferon-γ in Hallmark gene sets. The up-regulated gene sets in KEGG pathway included DNA replication, Proteasome, Mismatch repair, O-glycan biosynthesis, and Pentose phosphate pathway (Fig. 5D). Hence, our findings suggest that KPT-9274 regulates cell proliferation by suppressing the expression of these tumor growth-associated genes and pathways.
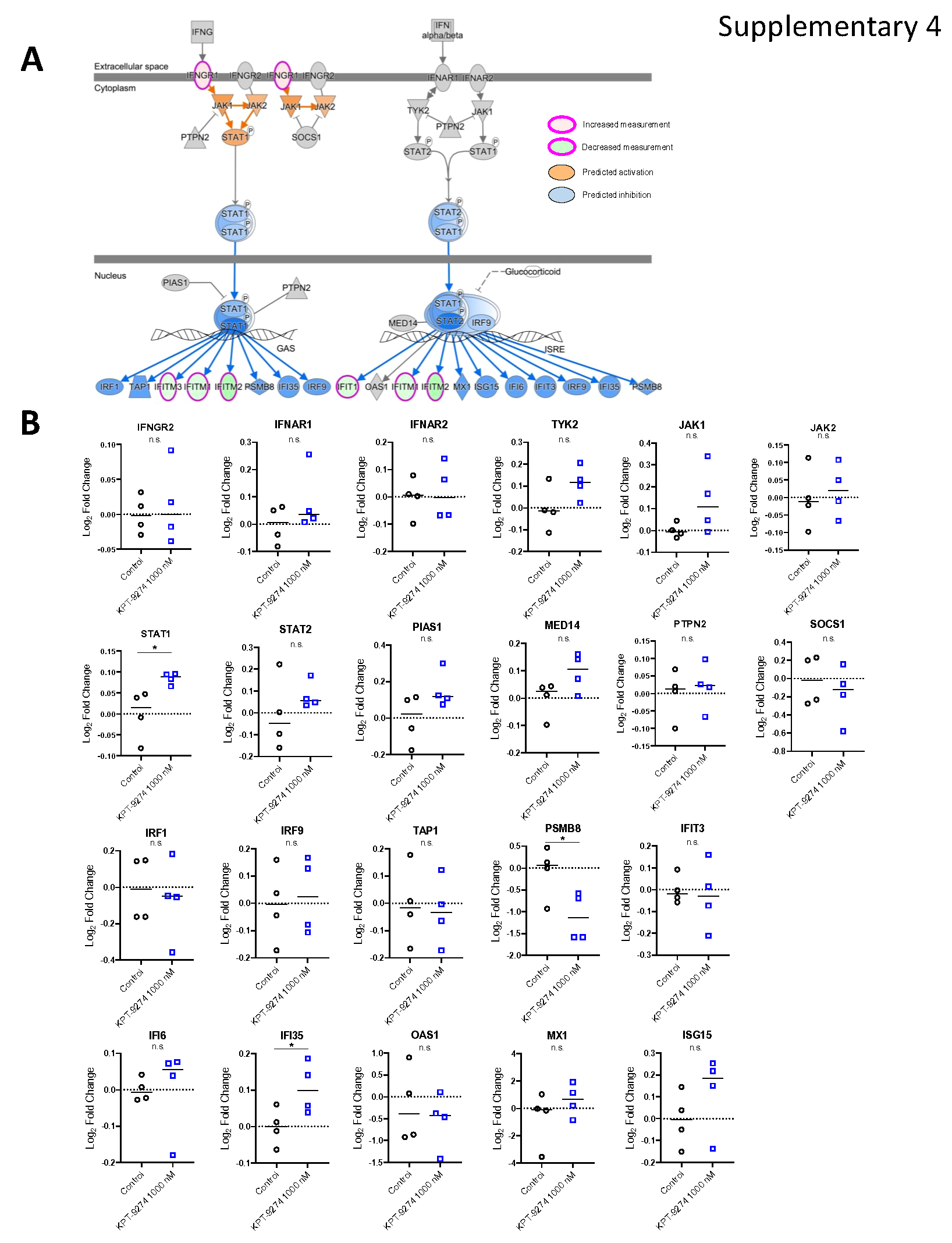
Ingenuity Pathway Analysis (IPA) unveiled changes in the Interferon signaling pathway, Remodeling of epithelial adherens junctions, and ILK signaling due to KPT-9274 treatment. The genes linked to Interferon signaling, namely IFNGR1, IFIT1, IFITM1, IFITM2, and IFITM3, showed varying expression patterns upon treatment (Supplementary Fig. 4A and 4B). Specifically, IFNGR1, encoding the IFN-γ receptor-1, was upregulated, while the others were downregulated (Fig. 5E). IFIT1 affects cancer cell behavior through Wnt/β-Catenin signaling [40], and IFITM1, IFITM2, and IFITM3 are related to antiviral functions [41]. To validate how these changes in transcriptomes affect protein expression, we tested expression of IFNGR1, IFIT1, IFITM1, IFITM2, and IFITM3 using Western blotting. IFITM1 was not detected (data not shown), and IFITM2/3 showed no significant differences between Control and KPT-9274 treatment. Interestingly, contrary to RNA-seq data, KPT-9274 significantly suppressed IFNGR1 expression, a membrane surface protein. Moreover, IFIT1 cytoplasmic expression was significantly down-regulated by KPT-9274 (Fig. 5F), suggesting that KPT-9274 downregulates Wnt/β-Catenin pathway via a suppression of IFNGR1 and IFIT1, contributing to the anti-tumor effects (Fig. 5G).
KPT-9274 down-regulates multiple kinase activities in the cytoplasm through a localization change of PAK4.
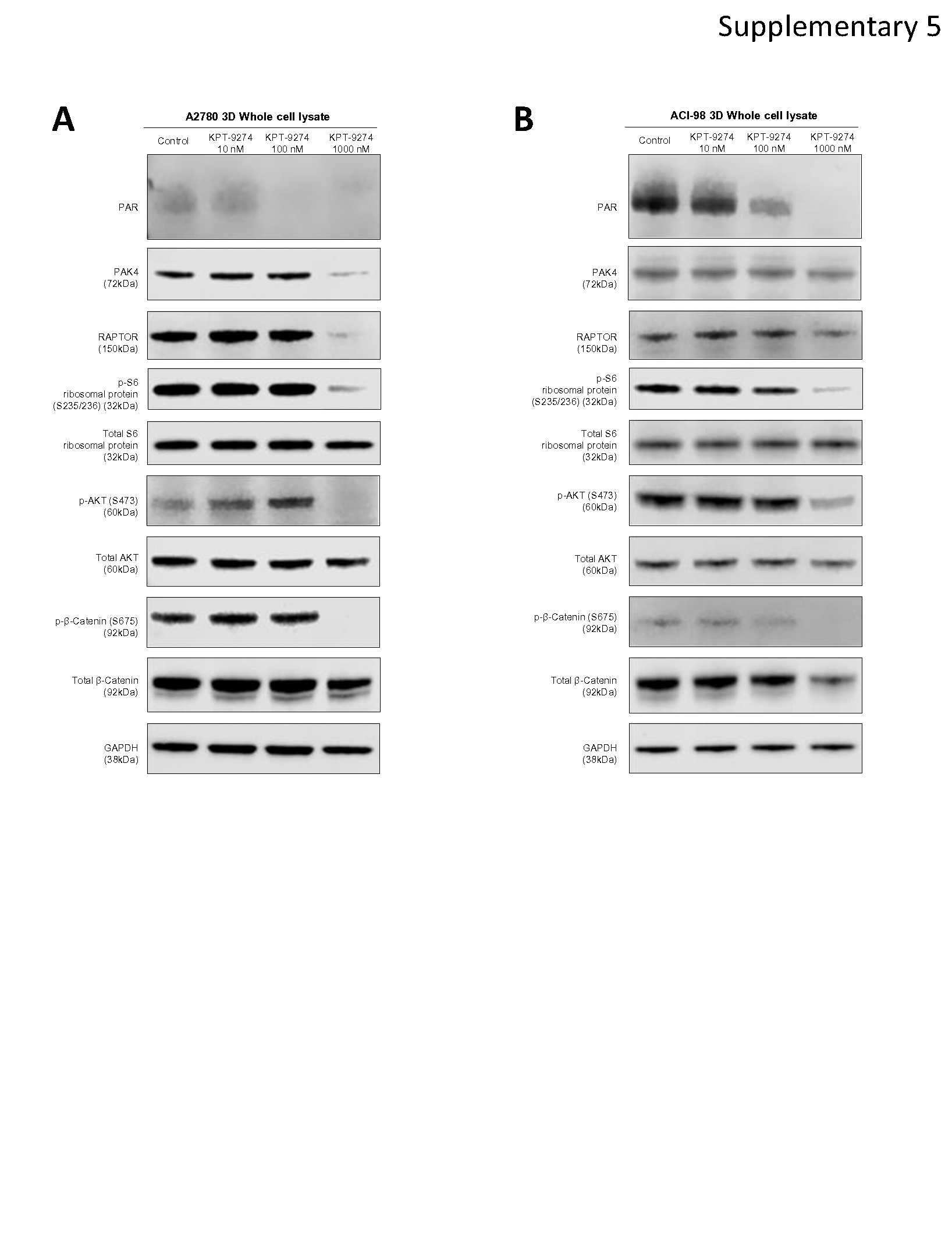
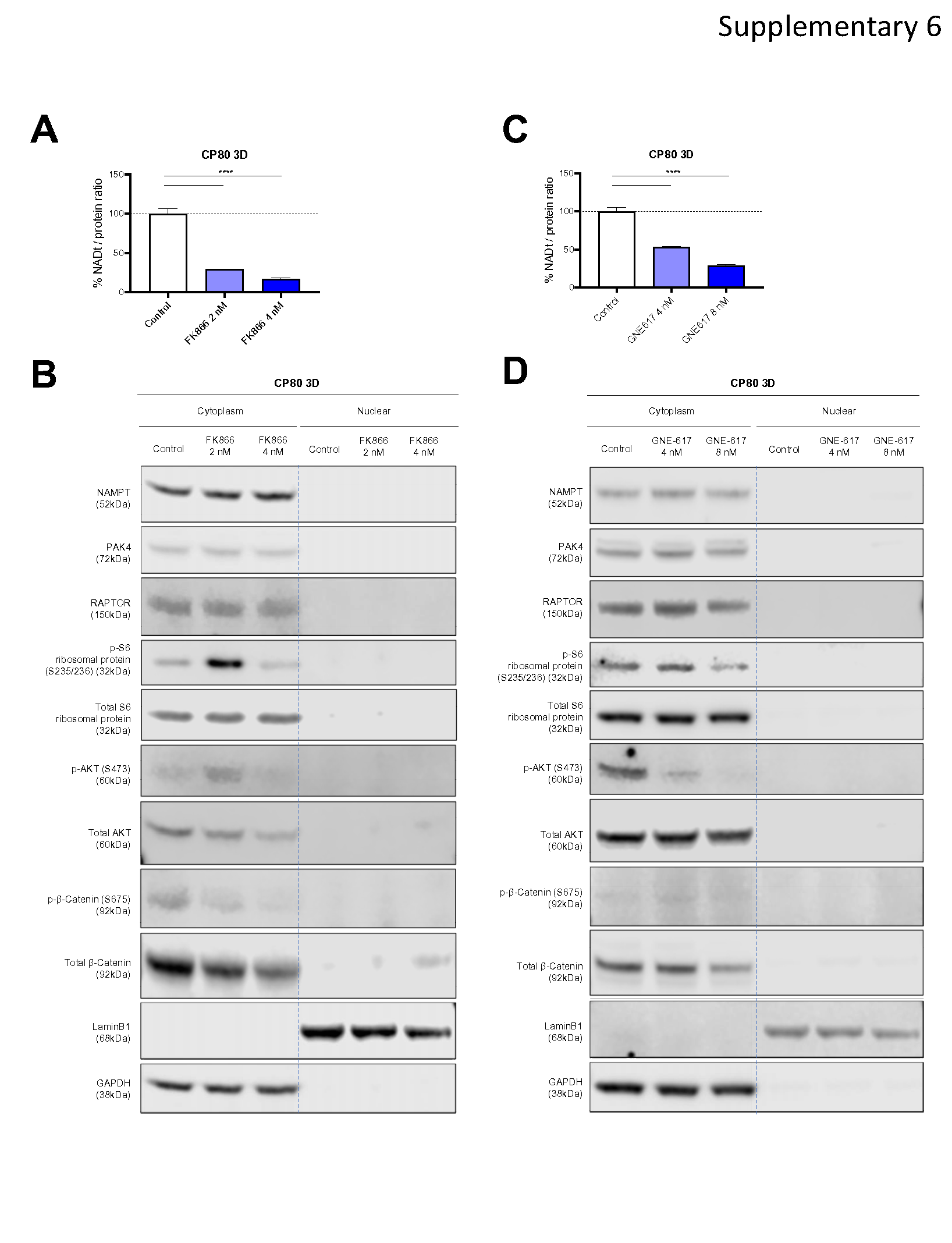
It has been shown that PAK4 regulates β-Catenin phosphorylation and mTOR complex function [19] [20] [21] [22] [23]. Hence, suppressing PAK4 leads to reduced kinase activity of various proteins, such as AKT, that are controlled by mTOR complexes. To validate the effect of KPT-9274 on kinase activity, we evaluated the expression of PAK4-affected proteins with Western blotting using cytoplasm and nuclear lysate. We also evaluated Poly (ADP-ribose) (PAR), which reflects the function of DNA repair [42], because RNA-seq results suggested KPT-9274 inhibited DNA repair. As expected, PAR expression was suppressed in both cytoplasm and nucleus following KPT-9274 treatment, suggesting DNA repair function was impaired by KPT-9274. Notably, PAK4, which was mostly localized in the cytoplasm before treatment, migrated into the nucleus after KPT-9274 treatment. In parallel to the shift of the localization of PAK4, cytoplasmic expression level of RAPTOR, Phospho-S6 Ribosomal Protein (Ser235/236), Phospho-AKT (Ser473), and Phospho-β-Catenin (Ser675) was decreased (Fig. 6A). RAPTOR and Phospho-S6 Ribosomal Protein (Ser235/236) reflect mTORC1 function [23]. Similar protein suppression was observed in 3D-cultured A2780 and ACI-98 cells, as well as CP80, in whole cell lysates (Supplementary Fig. 5A and 5B). Next, using FK866, the first-in-class NAMPT inhibitor, and GNE-617, a specific NAMPT inhibitor [43], we conducted a similar validation. Intriguingly, despite successfully inhibiting NAD+ production, the subcellular distribution of PAK4 remained unaltered with specific inhibition of NAMPT alone, while the impact on key proteins like RAPTOR, S6 Ribosomal Protein, AKT, and β-Catenin was inconsistent, displaying distinct patterns between NAMPT inhibitors (Supplementary Fig. 6A and 6B). These findings highlight that the alteration of PAK4 localization seems to be specific to KPT-9274.
In support of these Western blotting findings, immunofluorescence confocal imaging of spheroids also revealed the fluorescence intensity of Phospho-S6 Ribosomal Protein (Ser235/236) and Phospho-AKT (Ser473) in the 3D spheroids was suppressed with KPT-9274 treatment (Fig. 6B). Phospho-β-Catenin (Ser675) was difficult to detect (data not shown). To assess kinase activity from different perspectives, IncuCyte™ Kinase AKT Assay was next performed. AKT phosphorylation moves the green sensor from nucleus to cytoplasm. Conversely, AKT inhibition retains the sensor in the nucleus [44]. Interestingly, KPT-9274 treatment maintained the green signal in the nucleus, indicating suppressed AKT kinase activity (Fig. 6C). Nuclear Translocation Ratio, reflecting sensor movement [44], was reduced by KPT-9274 in a concentration-dependent manner, linked to inhibited cell proliferation (Fig. 6D). Overall, these findings suggested that KPT-9274 hindered cell proliferation by lowering cytoplasmic kinase activity through altering PAK4 localization (Fig. 6E).
The suppressed PAK4-mediated kinase activity by KPT-9274 treatment is NAD + -dependent.
To uncover whether the ability of KPT-9274 to suppress multiple kinase activities is a NAD+-dependent mechanism, we silenced NAMPT expression using siRNA. NAMPT-silenced cells showed approximately 60% less NAD+ content and about 75% less NAMPT expression corrected for GAPDH than control siRNA-treated cells (Fig. 7A and 7B). Adding NMN to the medium had no effect on NAMPT expression, while rescued total NAD to 80% of control. Consistent with the NAD+ production, PAR was suppressed in NAMPT-silenced cells and was rescued by NMN addition. However, NAMPT silencing did not impact PAK4, Phospho-S6 Ribosomal Protein (Ser235/236), Phospho-AKT (Ser473), and Phospho-β-Catenin (Ser675) (Fig. 7A and 7B). These findings suggest that solely reducing NAD+ through NAMPT silencing does not strongly suppress kinase activity.
Next, we tested whether supplemental NMN could rescue the kinase activity reduction caused by KPT-9274 treatment. As expected, KPT-9274 decreased NAD+ production by approximately 90% (Fig. 7C), while NAMPT expression increased (Fig. 7D), suggesting NAMPT upregulation due to NAD+ reduction. Importantly, NMN addition largely restored the suppressed PAK4, Phospho-S6 Ribosomal Protein (Ser235/236), Phospho-AKT (Ser473), and Phospho-β-Catenin (Ser675), indicating that suppressed PAK4-mediated kinase activity by KPT-9274 is NAD+-dependent (Fig. 7C and 7D). In conclusion, KPT-9274 demonstrated a promising activity against NAMPT or PAK4-driven cancer growth, suggesting it is a potential novel treatment for platinum-resistant ovarian cancer.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}