Social behavior is inextricably linked to the immune system. Although IFN-γ is known to be involved in social behavior, yet whether and how it encodes social memory remains unclear. In this study we demonstrated that IFN-γ injection induces the priming of hippocampal microglia by activating STAT1 pathway, thereby disrupts hippocampal microglia-neuron interactions and neural circuit link to social memory, resulting impairment in social memory. Blocking STAT1 pathway or inhibiting microglial priming may be strategies to reduce the effects of IFN-γ on social behavior. To our knowledge, this study is the first to clarify in detail the properties of IFN-γ-primed microglia and their effects of on social behavior and the underlying mechanisms.
Although at first glance inflammation and social behavior may appear unrelated, research points to an important role for inflammation in shaping social processes [32]. It is intriguing that IFN-γ, predominately thought of as an anti-pathogen cytokine, can play such a profound role in maintaining proper social function [13]. In the current study, we found the mice injected with 500 ng of IFN-γ showed significant decrease in social index and social novelty recognition index, suggesting that overweight IFN-γ in brain can cause social avoidance and impairment in social recognition memory. It also had been reported that there were significant negative correlations between IFN-γ and cognition in patients with schizophrenia [33] or positive correlations between IFN-γ and social impairment in patients with autism spectrum disorder [34]. These findings could partly explain why human and animals adopt social avoidance when severely infected with a virus [35-37]. It may be that the upregulated IFN-γ in brain can reduce the likeliness of spreading pathogens by reducing social interaction. Studies similar to the IFN-γ-mediated impairment of social memory found in this study have been reported that T cell-derived IFN-γ signaling in microglia underlies spatial-learning defects via virus-target-specific mechanisms [38]. Interestingly, the effects of IFN-γ knockout on cognitive function and social behavior were opposite in mice. In one study, mice deficient in IFN-γ exhibit social deficits and hyper-connectivity of front-cortical brain regions [13]. In contrast, another study found that IFN-γ deficiency led to enhanced performance on cognitive tasks [25]. Combined with our findings in the effect of excessive IFN-γ on sociability and social memory of male mice in present study, IFN-γ may differentially impact the social vs. non-social domains, and of course, these relationships may also be altered by its concentration in brain and interactions with other biopsychosocial factors.
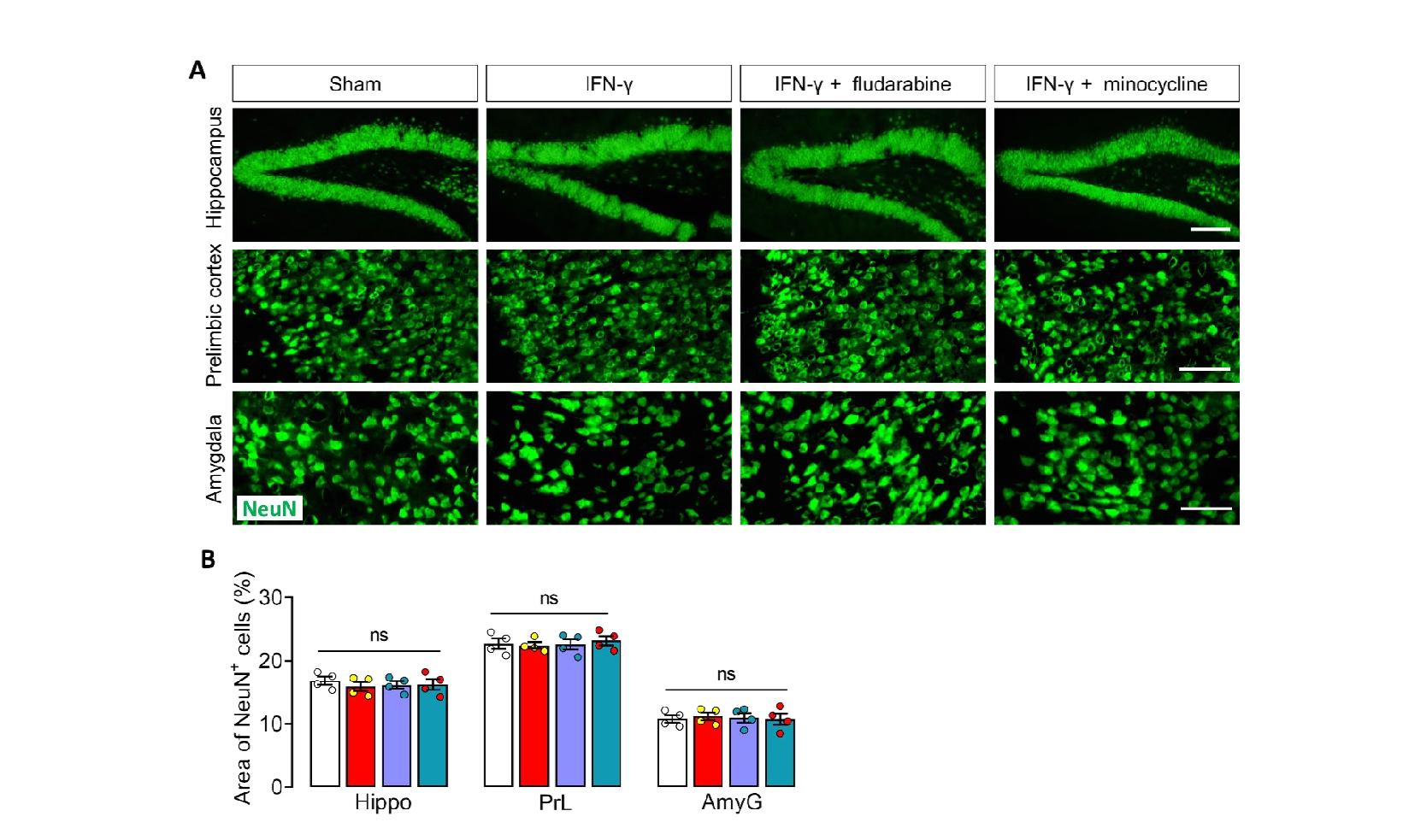
Our results reveal a novel role for IFN-γ mediated neuro-immunity response in regulating social behaviors. These changes in social behavior as a result of pro-inflammatory cytokine activity provided the first clues that inflammation may play an important role in shaping social behavior. IFN-γ signaling as a selective key regulator of immune cell trafficking across the choroid plexus epithelium, which plays an important role in central nervous system (CNS) immune surveillance and neuroimmunomodulatory [20]. IFN-γ priming induces substantial proliferation and moderate activation of microglia that is capable of slowing neural information processing [24]. This mechanism might contribute to social dysfunction in brain disease featuring elevated IFN-γ levels, blood–brain barrier leakage, and/or T cell infiltration. Hippocampus, prelimbic cortex and amygdala are considered to be the major brain region involved in the regulation of social behavior [39-43]. Changes in microglia morphology and function involves in social behavioral defects [44, 45]. Interestingly, our data showed that IFN-γ-injected samples showed increased density and area of Iba1+ cells and decrease in length of microglial branches in the hippocampus but not in prelimbic cortex and amygdala. These results suggested that IFN-γ specifically induces microglial priming in hippocampus. This may be due to the location of hippocampus in regards to the lateral ventricles, where the IFN-γ was introduced into the CNS, making them more susceptible to the local environment [21]. In addition, microglia in the hippocampus showed higher activation levels than the cortex and were characterized by larger cell bodies, fewer branches. A more intriguing hypothesis is that the hippocampus provides a protected neurogenic niche and the resident microglia are more sensitive to stimulation.
Microglia priming is often implicated by the onset of inflammation and increase of immune response activity, which associates with a variety of cellular adaptations, including changes in morphology, up-regulation of receptors, and increased levels of proinflammatory cytokines [21, 23, 24]. In this study, we found that IFN-γ-primed microglia is characterized by increase of CD68, CD86, CD74, CD11b, CD11c, CD47, IL-33, IL-1β, IL-6 and iNOS expression, but decrease of MCR1, Arg-1, IGF-1 and BDNF in hippocampus. We also found that microglia in IFN-γ-injected mice were more susceptible to immune stimulation in the DG than microglia in Sham animals. LPS administration led to more severe disease-like symptoms as well as anxiety-like behaviors in IFN-γ-injected mice. LPS administration also led to more extensive microglial priming, greater areas of Iba1+ staining, shorter branches of microglia, and higher concentrations of IL-1β in the hippocampus of IFN-γ-injected mice than in Sham animals. These results showed that IFN-γ can prime exaggerated microglial responses upon stimulation with microbial or endogenous ligands, such as bacterial LPS. Minocycline is a semi-synthetic, second-generation tetracycline analog which is effectively crossing the blood-brain barrier, effective against gram-positive and -negative infections [46]. As the inhibitor or phenotypic modulators of microglia, minocycline has been reported to exert neuroprotective effects over various experimental models or patients involved microglial activation such as cerebral ischemia [47], traumatic brain injury [48], depression [49], social stress [50], cognitive deficit [51] and so on. In present study, we found that minocycline treatment prevented microglial priming and rescued abnormal crosstalk between microglia and neuron in hippocampus of IFN-γ-injected mice, as well as improved social memory. These results suggest that inhibition of microglial activation using minocycline has potential development prospects for social defects caused by immune over-activation.
Deficient neuron-microglia signaling results in impaired functional brain connectivity and social behavior has been reported [52-54]. The continuous crosstalk between microglia and neurons is required for microglia housekeeping functions and contributes to brain homeostasis, through which CX3CL1/CX3CR1 and CD200/ CD200R axis [55-57]. Study showed that microglia and its receptors CX3CR1 are considered the key contributors in regulation of social behavioral adaptation [58] Social isolation caused deficits in the prepulse inhibition in the WT mice but not in the CX3CR1 knockout mice [59, 60], suggesting that the function of microglia regulated by CX3CR1 might participate in social behaviors. In addition, lack of CX3CL1/ CX3CR1 signaling led to reduced responsiveness to social defeat in mice, suggesting CX3CL1/CX3CR1 signaling is needed to social response [61]. Mice lacking the chemokine receptor CX3CR1 exhibit a consequent deficit in synaptic pruning, decreased functional brain connectivity, deficits in social interaction that have been previously associated with autism and other neurodevelopmental and neuropsychiatric disorders [52]. Consistent with these reports, in this study, we found IFN-γ injection reduced hippocampal levels of CX3CL1 and its receptor CX3CR1, and increased expression of CD200R but did not affect its ligand CD200 expression, which are the intercommunicating molecules between neuron and microglia. The alteration of neuron-microglia communication contributes to brain disease states with abnormal synaptic function [52, 53]. In this study, we found the PSD-95, a marker of excitatory postsynaptic membrane, which swallowed into the microglia were significantly increase in hippocampus of IFN-γ-injected mice, as well as fewer postsynaptic clusters of PSD95and synapsin than Sham mice. The hippocampal levels of glutamate transport-related receptor GluA2 was reduced in compared with Sham mice. Another interesting finding in this study was that although microglia in the prelimbic cortex and amygdala were not primed by IFN-γ as well as the unaltered NeuN+ cells, the levels of GluA2 were upregulated in these two regions, which is the opposite of what happens in the hippocampus. It had been found that IFN-γ priming induces substantial proliferation and moderate activation of microglia that is capable of slowing neural information processing [24]. Combine the literature with our findings reveal that IFN-γ-primed microglia in hippocampus can communicate abnormally with neurons, which disrupts synaptic function of hippocampal neurons and causes changes in neural circuit link to social behaviors.
Since IFN-γ mediated innate immune response is initiated by interferon regulatory factors which are regulated by phosphorylated signal transducer and activator of transcription 1 (STAT1) [62], the role of JAK/STAT1 signal transduction in social behavior is widely noticed [13, 63, 64]. Meta-analysis of the transcriptomes of a range of organisms reveals that rodents, fish, and flies elevate IFN-γ/JAK-STAT1-dependent gene signatures in a social context, suggesting that the IFN-γ signalling pathway could mediate a co-evolutionary link between social/aggregation behavior and an efficient anti-pathogen response [13]. In this study, we found that IFN-γ treatment increased the levels of pSTAT1 and nuclear translocation of STAT1 in microglia. STAT1 inhibitor fludarabin treatment suppressed microglial priming in hippocampus of IFN-γ-injected mice, prevents the dysregulation of glutamate transport by rebalanced the intercommunicating molecules between neuron and microglia. Of course, the IFN-γ-induced social dysfunction was also alleviated parallelly by fludarabin. These results suggested that STAT1-mediated priming of hippocampal microglia play a key role in social dysfunction of IFN-γ-injected mice.
{kind=link}