A PapB/PefB paralogue on pMB2 interferes with autoaggregation. pMB2 was originally extracted from a commensal Escherichia isolate from Nigeria [20, 21]. In the course of functional analyses of fitness loci [6], we additionally observed that DH5α carrying the pRMKO miniplasmid autoaggregated significantly, particularly under low nutrient conditions. As shown in Figure 1a, autoaggregation in liquid media was prominent in DH5α (pRMKO) but absent in DH5α (pMB2) cultures grown under the same conditions (p<0.001). Cloning the 32 Kb deleted region into pBluescript also reduced autoaggregation of DH5α compared to an isogenic strain carrying the vector alone (Figure 1a). We examined the sequence of the region deleted from pRMKO for genes with the potential to contribute to autoaggregation. The best candidate is small open reading frame (orf) located from 11,258-11,530 of the plasmid. The predicted amino acid sequence of this gene was 29% identical/ 50% similar to E. coli PapB from uropathogenic strain J96 (Genbank Accession ELL39276.1) and was 32% identical/ 48% similar to PefB, a PapB paralogue from Salmonella enterica serovariety Typhimurium strain LT2 (Genbank Accession AAL23523). There are no fimbrial genes proximal to the pMB2 papB-like gene, however there are mobility genes on either side and the papB-like gene region has a noticeably lower G+C content than the surrounding region (Figure 1b).
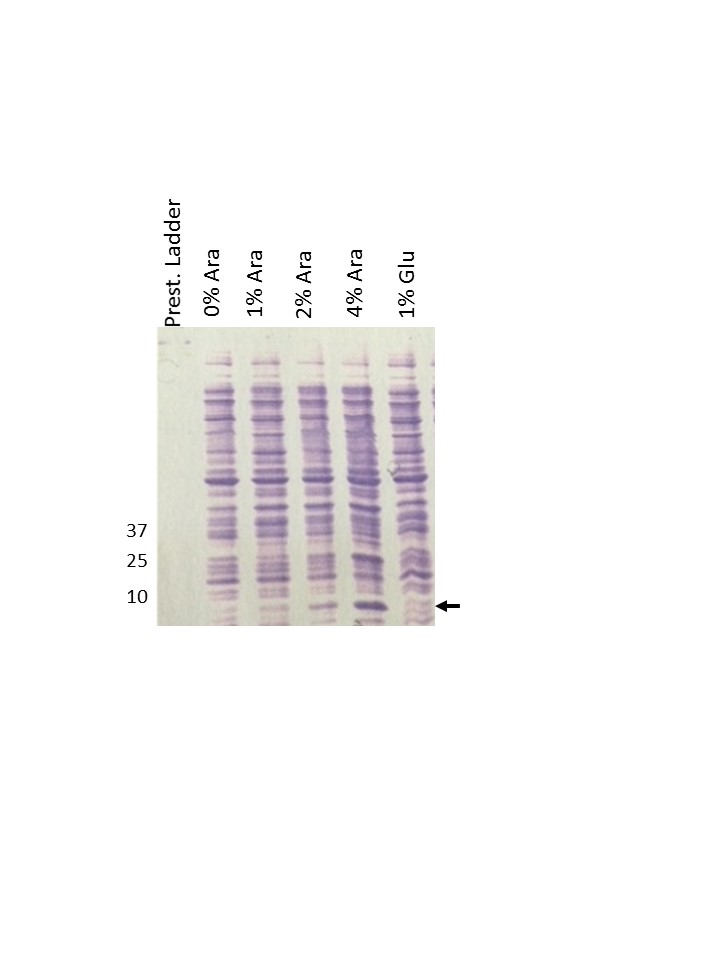
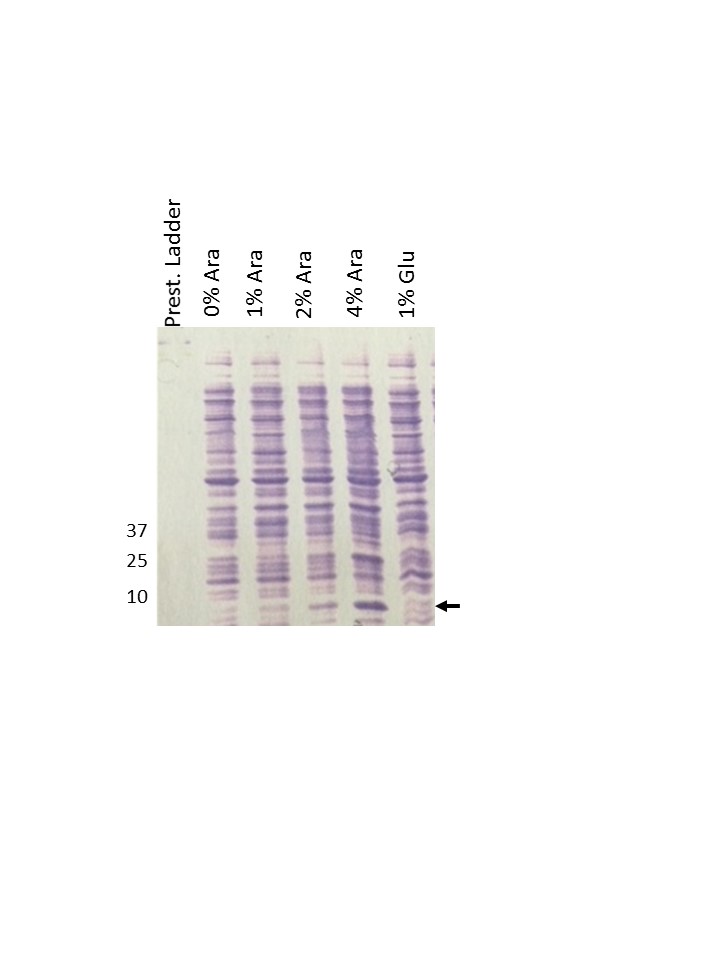
We cloned the papB/pefB paralog under the control of the arabinose promoter and measured the clone’s ability to complement pRMKO hyperautoaggregation. As shown in Figure 1c, when the papB/pefB paralogue is induced by arabinose, autoaggregation was diminished in DH5α (pRMKO, pINKPefB). This phenotype was not seen when the arabinose promoter is glucose-repressed (Figure 1c). Based on this finding, we attribute the hyperautoaggregation phenotype conferred by pRMKO to deletion of the pMB2 papB/pefB paralogue.
The pMB2 papB/pefB paralogue acts on chromosomally-encoded effectors
PapB binds between papI and papB in the extraintestinal pathogenic E. coli pap cluster as well as, if less strongly, upstream of fimE [10]. R61A and C65A substitutions, within a hydrophilic region of PapB, produce derivatives unable to bind DNA [11]. As shown in Figure 2, pMB2 PefB paralogue has a cysteine at the position equivalent to 65 in PapB. It does not have an arginine at position 61 but does have a positively-charged lysine residue at the equivalent position. Positions 86 and 91 at the PapB carboxy terminus are also essential for switching activity [8]. Holden et al reported that L82F or F83Q substitutions, which altered these PapB residues to the equivalents in the non-binding DaaA paralogue, reduced switching. When both substitutions were made, activity was obliterated [8]. As shown in Figure 2, the pMB2 PefB paralogue has a leucine at the position equivalent to PapB 82 but has an asparagine substitution at position 83, and so, even though it is considerably dissimilar to PapB, it could conceivably inhibit fim switching. However, Figure 2 illustrates that there are many sequence differences between the allele we have identified and previously studied members of this family making it impossible to infer the nature and degree of its function or activity from sequence alone. In DH5α (pMB2), PapB/PefB could be repressing either chromosomally-encoded fim genes or pMB2 factors not deleted from pRMKO. From the complete sequence of pMB2, we did not identify genes encoding chaperone-user fimbriae or any other fimbrial type other than the F (conjugative)-pili, which are known to promote autoaggregation [6, 22, 23], although they lack a type I fimbriae-like switch. We additionally acknowledge that any of the several pMB2 orfs of unknown function could contribute to autoaggregation. To determine which possibility could be at play, we transformed pMB2 and pRMKO into ORN172, a fim-negative E. coli strain [24]. In this Dfim background, there were no papB/pefB-associated differences in autoaggregation (Figure 1d). Therefore, while it remains to be confirmed in direct experiments, this finding suggests that interactions of papB/pefB with core chromosomal factors, likely fimbriae, account for the hyperautoaggregation seen in DH5α (pRMKO) and that the hydrophobic residue at position 83 of PapB, may be less critical for FimB regulation than the presence of a leucine at position 82.
PapB is not known to directly regulate conjugative pili, but a fim repressor could conceivably enhance F-pilus-mediated autoaggregation and conjugation by preventing occlusion by chromosomally-encoded fimbriae. We performed an initial test of this hypothesis by comparing conjugative transfer rates from DH5α (pMB2) and DH5α (pRMKO) to strain EC1502. (The pRMKO miniplasmid contains the complete conjugative gene region of parental pMB2 plasmid). We recorded a transfer rate of 7.9 X 10-6 for DH5α (pMB2) in solid matings, compared to 3 X 10-5 of control plasmid pMB80 [25]. In contrast, conjugation from DH5α (pRMKO) could not be detected at the limits of the assay. Due to the low conjugation rate overall and the impossibility of detecting rates below 1 X 10-8 we could not perform a robust complementation experiment. However, altogether the available data support the idea that papB/pefB repressed genes could interfere with pMB2 conjugative transfer.
The pMB2 papB/pefB paralogue is widely distributed and largely overlooked on extraintestinal E. coli plasmids
A BLAST search of the pMB2 nucleotide sequence range 7247-14962, representing the papB/ pefB paralogue, its flanking low G+C region and the adjacent NikM/Hmu genes, reveal that it is a common plasmid-borne feature. Importantly, plasmids of E. coli O25b:H4-ST131 strains, commonly implicated in bloodstream, urinary and other extraintestinal infections, bear the entire region depicted in Figure 1b. On the E. coli str. UMN026 plasmid p1ESCUM, often used an extraintestinal E. coli reference sequence (GenBank Accession Number: CU928148.1) [26], the entire region is present, but the papB/pefB paralog is not annotated as is the case for some well-known resistance plasmids [27], but not for others.
Cystitis and pyelonephritis isolates are more likely to contain one or more pap operons than other E. coli. Holden et al (2006) [28], who observed this, also showed that cystitis and pyelonephritis isolates are better able to agglutinate red blood cells and attributed this to a higher copy-number and activity of intracellular PapB, which promoted expression of alternate fimbriae whilst shutting type I fimbriae OFF. papB/pefB paralogues on plasmids could achieve this effect irrespective of the presence of a full fimbrial operon consequently modulating the expression of core adhesins for in vivo adherence and/or exposure to the immune system. Thus, virulence factor profiling of pathogenic E. coli needs to take into account the available fimbrial genes as well as fimbrial regulators some of which—like the gene we characterize—could be plasmid-borne. As our papB/pefB paralogue is borne on a plasmid conferring resistance to no less than eight antimicrobial classes, antibiotic selective pressure could ultimately alter colonization and virulence profiles of strains in the wild.
In conclusion, pMB2 is self-transmissible by an F-type conjugative system that may function more effectively due to repression of chromosomal adhesins by a papB/pefB paralogue located on the same plasmid. Plasmids almost identical to pMB2 have been isolated from the USA (Accession number CP054458), Colombia (Acession number CP010372), France (Accession number LO017738), the UK (Accession number CP023372, CP018990) and elsewhere. The papB/pefB paralogue we describe is highly conserved among them and therefore the autoaggregation it represses is likely common, warranting further study. Our findings add to experimentally validated reports of two other non-resistance loci of pMB2 that promote E. coli survival and adaptability [6, 29]. Thus there are multiple genes resistance plasmids hat compensate the costs of their carriage or, as in this case, potentially propagate transmission. This study also provides yet another example [30] horizontally acquired DNA that radically affects the expression of recipient cell core genes.
{kind=link}
{kind=link}