Equol production by LAB and bifidobacteria strains harbouring dzrP, ddr-tdrP and ifcAP
The work previously developed by our group [12] was extended with new LAB and bifidobacteria strains, such as S. thermophilus INIA 461, E. faecalis INIA 4, L. reuteri INIA P572, L. fermentum INIA P143, L. paracasei INIA P272, L. rhamnosus INIA P540 and B. breve INIA P734. Moreover, we also used L. lactis MG1363, L. fermentum INIA 584L and L. fermentum INIA 832L previously used by Ruiz de la Bastida et al. (2021). We observed that the new strains harbouring dzrP and ddr-tdrP either did not produce equol from daidzein, or the equol production was in concentrations lower than that shown by L. fermentum INIA 584L and L. fermentum INIA 832L, and even lower than those produced by L. lactis MG1363 (Figure 1A). We did not observe equol production from daidzein by B. breve INIA P734, S. thermophilus INIA 461, L. rhamnosus INIA P540 or E. faecalis INIA 4 harbouring dzrP and ddr-tdrP. However, L. reuteri INIA P572, L. fermentum INIA P143 and L. paracasei INIA P272 harbouring dzrP and ddr-tdrP produced between 4.76 ± 0.94 and 16.34 ± 1.90 µM of equol from daidzein (Figure 1A).
The incorporation of the heterologous expression of dihydrodaidzein racemase (ifcA) only showed an increase in equol production in L. fermentum INIA 584L and L. fermentum INIA 832L. The production of equol from daidzein by L. fermentum INIA 584L increased from 44.34 ± 3.26 to 172.70 ± 10.02 µM with the incorporation of the heterologous expression of ifcA. Meanwhile, the equol production from daidzein by L. fermentum INIA 832L increased from 38.34 ± 6.45 to 165.70 ± 12.41 µM.
All the new strains harbouring tdr-ddrP showed equol production from DHD, although at concentrations always lower than 25 µM. The production of equol from DHD for these strains was much lower than the production of equol using L. fermentum INIA 584L and L. fermentum INIA 832L (Figure 1B). The incorporation of the heterologous expression of ifcA (ifcAP) did not show an increase in equol production from DHD in any of the strains used.
Effect of the heterologous expression of ifcA in the production of equol from daidzein and DHD
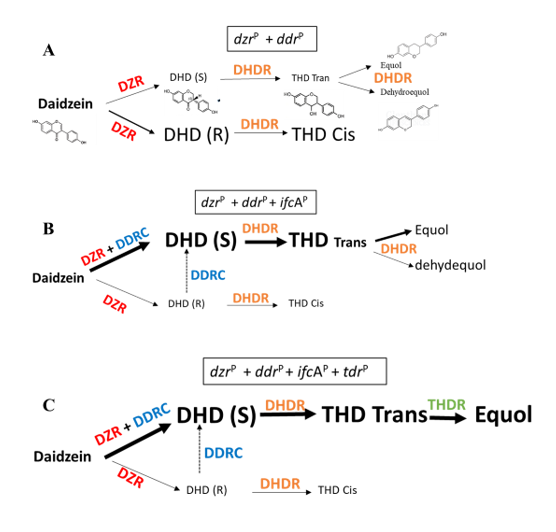
After the results of the equol production by LAB and bifidobacteria strains observed in this work and previously by Ruiz de la Bastida et al. [12], we focused our work on the heterologous expression of ddr and ifcA, because both DDRC and DHDR could be responsible for the differences in the equol production observed between the different bacteria studied.
L. lactis strains harbouring (dzrP + ddr-tdrP) and (dzrP + ddr-tdrP + ifcAP) showed an equol production of 22.83 ± 2.67 µM and 20.76 ± 2.26 µM, respectively. Hence, the heterologous expression of ifcA in L. lactis MG1363 did not seem to affect the equol production. Whereas, as mentioned above, the heterologous expression of ifcA together with dr and ddr-tdr (dzrP + ddr-tdrP + ifcAP), in L. fermentum INIA 584L and L. fermentum INIA 832L, showed an increase in equol production 4 to 5 times greater with respect to that observed without the presence of ifcA (Table 2).
The most interesting data appeared when L. lactis harbouring dzrP and tdr-ddrP (dr + ddr-tdr) were co-incubated with L. fermentum INIA 584L or L. fermentum INIA 832L harbouring ifcAP (dzrP + ddr-tdrP + ifcAP L. fer.584L or ifcAP L. fer.832L). Equol production increased two to three-fold in the presence of the heterologous expression of ifcA in L. fermentum INIA 584L (42.15 ± 1.67 µM) and L. fermentum INIA 832L (43.25 ± 3.18 µM) (Table 2). This data demonstrated that the heterologous expression of ifcA from S. isoflavoniconvertens in L. fermentum strains helps to increase equol production by L. lactis harbouring dzrP and tdr-ddrP.
On the other hand, the coincubation of L. lactis harbouring dzrP and L. fermentum 584L harbouring tdr-ddrP showed a production of 36.56 ± 6.34 µM of equol, and the addition of L. fermentum INIA 584L harbouring ifcAP showed an important increase in equol production (158.32 ± 13.12 µM). Equol production was similar when L. fermentum 584L harbouring tdr-ddrP and ifcAP were combined with L. lactis harbouring dzrP (158.32 ± 13.12 µM) and L. fermentum 584L harbouring dzrP (172.70 ± 10.02 µM) (Table 2).
Finally, the heterologous expression of dr, tdr-ddr and ifcA in L. fermentum INIA 584L and L. fermentum INIA 832L showed a higher production of equol from daidzein than from DHD, whereas L. lactis harbouring tdr-ddrP showed higher equol production from DHD (Table 3). The heterologous expression of ifcA did not demonstrate any effect on equol production from DHD in L. lactis and L. fermentum strains. However, the combination of L. lactis harbouring tdr-ddrP and L. fermentum INIA 584L harbouring ifcAP did show an effect of the heterologous expression of ifcA, and a significant increase in equol production from DHD was observed.
Effect of the heterologous expression of ifcA on DHD production from daidzein
In order to understand the differences observed in the amount of equol produced by the presence of DDRC, we studied how DDRC influenced the production of DHD and THD cis and THD trans. As mentioned above, the effect of DDRC on the production of equol from DHD was low, and, in addition, we observed that the heterologous expression of (dzrP + tdr-ddrP + ifcAP) was capable of transforming most of the daidzein into equol in L. fermentum INIA 584L and 832L. Thus, something is happening in the transformation of daidzein into DHD and THD in L. fermentum INIA 584L and 832L that facilitates the production of equol when DDRC is present.
The heterologous expression of ifcA in L. fermentum INIA 584L produced an increase in the transformation of daidzein in DHD by L. fermentum INIA 584L dzrP following 4, 8, 12, 16 and 24 hours of incubation (Figure 2A). At longer times, 36 hours or more, we did not observe a significant effect of ifcA on the production of DHD, since most of the daidzein was transformed into DHD. We could not differentiate DHD-R from DHD-S. However, we observed an increase in the rate of transformation of daidzein into DHD with the heterologous expression of ifcA in L. fermentum strains.
On the other hand, no difference was observed in DHD production when L. lactis harbouring dzrP or L. lactis harbouring dzrP and ifcAP were incubated with daidzein (Figure 2B). However, when L. lactis dzrP was incubated with L. fermentum INIA 584L ifcAP, we observed an increase in the rate of transformation of daidzein into DHD, similar to that observed in Figure 2A.
Effect of the heterologous expression of ifcA on the production of THD from daidzein
ddr was cloned separately from tdr to observe the formation of THD cis and THD trans in order to understand the differences observed in the amount of equol produced by the presence of DDRC. The incubation of L. lactis and L. fermentum INIA 584L and L. fermentum INIA 832L harbouring dzrP and ddrP with daidzein clearly showed the formation of both THD cis and THD trans. The absence of the heterologous expression of ifcA, L. fermentum INIA 584L and L. fermentum INIA 832L showed that the area ratios of THDcis/THDtrans measured by HPLC-ESI/MS were 1.51 ± 0.14 and 1.48 ± 0.21, respectively (Table 4). While an area ratio of 1.26 ± 0.08 was observed in the case of L. Lactis. This demonstrated a higher THD cis production from daidzein in the absence of DDRC.
The heterologous expression of ifcA together with dzr and ddr (drP + ddrP + icfAP) clearly showed a change in the ratio of THD trans/THD cis areas as a consequence of an increase in THD trans and a decrease in THD cis concentrations. Thus, the concentration of THD trans produced in the presence of the DDRC was three times that of THD cis in L. fermentum INIA 584L and double that in L. fermentum INIA 832L.
L. lactis MG1363 (drP + ddrP) showed a THD trans/THD cis ratio of 1.26 ± 0.08, whereas the incorporation of the heterologous expression of ifcA (drP + ddrP + icfAP) showed a ratio of 1.18 ± 0.06. When L. lactis MG1363 (drP + ddrP) was incubated together with L. fermentum INIA 584L ifcAP, it showed a ratio of 0.77 ± 0.11. A higher concentration of THD trans compared to THD cis was also observed with the heterologous expression of ifcA, and this difference increased with the heterologous expression of ifcA in the L. fermentum strains (Table 4).
L. fermentum strains (drP + ddrP) showed a THD production between 10 and 20 times greater than that shown by L. lactis (drP + ddrP), both in the presence and absence of the heterologous expression of ifcA (Figure 3A). These differences were observed despite the fact that most of the daidzein had been transformed into DHD in both the L. fermentum strains and in L. lactis. This data demonstrated a much higher activity of DHDR in these L. fermentum strains.
In addition, the heterologous expression of dzr and ddr (drP + ddrP) in both L. fermentum strains showed the production of equol and a compound that was identified as dehydroequol (C15H12O3) (Figure 3B). Moreover, the heterologous expression of ifcA together with that of dzr and ddr increased the production of equol and dehydroequol. While the heterologous expression of ddr in L. lactis did not show the production of these compounds, possibly as a consequence of the lower production of THD.
Effect of the heterologous expression of ifcA on the production of THD from DHD
The heterologous expression of ddr in both L. fermentum strains showed a 5 to 10 fold greater production of THD from DHD compared to when ddr was expressed in L. lactis (Figure 3C). As for daidzein, THD production was higher in the L. fermentum strains, demonstrating the highest DHDR activity in the L. fermentum strains. On the other hand, the heterologous expression of ifcA did not show variations in the THD cis/THD trans ratio in the transformation of DHD in THD (Table 4).
The heterologous expression of ddr in L. fermentum INIA 584L and L. fermentum INIA 832L also showed equol (32.18 ± 3.35 and 29.92 ± 5.76 µM) and dehydroequol (8.18 ± 2.45 and 9.06 ± 1.85 µM) production from DHD. From DHD, we also observed the production of equol (5.19 ± 2.32 µM) and dehydroequol (3,67 ± 1,63 µM) with the heterologous expression of ddr in L. lactis, although at a significantly lower concentration than that produced in the L. fermentum strains (Table 4). Moreover, the heterologous expression of ifcA in L. fermentum INIA 584L INIA 832L showed an increase in equol production from DHD by DHDR, although the increase in equol production due to the presence of DDRC was not significant (Table 4).
Role of the heterologous expression of tdr in equol production:
tdr was also cloned independently of ddr and its heterologous expression was analyzed. As expected, coincubations of the different strains harbouring ddrP with the same strains harbouring tdrP showed an increase in equol production mainly by the L. fermentum strains. So, whereas L. fermentum INIA 584L and 832L (drP + ddrP + ifcAP) produced 44.13 ± 5.63 and 45.56 ± 7.80 µM of equol from daidzein, respectively, the incorporation of the heterologous expression of tdr (drP + ddrP +tdrP + ifcAP) showed an equol production of 141.14 ± 8.23 and 124.27 ± 8.34 µM, respectively (Table 5). The incorporation of tdrP produced a decrease in both THD cis and THD trans as a consequence of the increase in the transformation of THD into equol. Moreover, when we used DDRC in addition to DHDR and THDR the concentrations of both THD cis and THD trans practically disappeared (Figure 6).
As mentioned above, DHDR acts on DHD to mainly produce THD, and also to produce equol and dehydroequol. However, the combination of DHDR and THDR increases the efficiency of equol production and decreases the concentration of dehydroequol (Table 4).
The heterologous expression of ddr and tdr in the same cells (ddr-tdrP), or in different cells (ddrP + tdrP), showed differences in equol production (Table 3). Coincubation of tdr and ddr on separate cells (tdrP + ddrP) showed a decrease in equol production. We observed that L. fermentum INIA 584L and L. fermentum INIA 832L (drP+ ddr-tdrP+ ifcAP) produced 172.70 ± 10.02 and 165.70 ± 12.41 µM of equol, respectively, from daidzein, whereas L. fermentum INIA 584L and L. fermentum INIA 832L (drP + ddrP + tdrP + ifcAP), produced 141.14 ± 8.23 and 124.27 ± 8.34 µM of equol, respectively (Table 4). Similar results were observed in the equol production from DHD, although the differences were not significant.
Effect of NADPH on equol production
To study the influence of the presence of NADPH on equol production, L. lactis MG1363 and L. rhamnosus INIA P540 harbouring (drP + ddr-tdrP + ifcAP) or (ddr-tdrP + ifcAP) were incubated with daidzein and DHD, respectively, in the presence and absence of NADPH. The presence of NADPH showed a significant increase in equol production from both daidzein and DHD. L. lactis MG1363 harbouring (drP + ddr-tdrP + ifcAP) and (ddr-tdrP + ifcAP) produced 20.26 + 1.20 and 29.50 + 2.16 µM of equol from daidzein and DHD, respectively, in the absence of NADPH. Whereas the presence of NADPH showed an equol production of 29.52 + 2.95 and 38.27 + 1.98 µM from daidzein and DHD, respectively (Figure 4).
On the other hand, L. rhamnosus INIA P540 (drP + ddr-tdrP + ifcAP) was unable to produce equol from daidzein, and L. rhamnosus INIA P540 (ddr-tdrP + ifcAP) was only able to produce 1.55 ± 0.85 µM of equol from DHD, in the absence of NADPH. Whereas, L. rhamnosus INIA P540 (drP + ddr-tdrP + ifcAP) produced 8.05 ± 1.10 µM of equol from daidzein, and L. rhamnosus INIA P540 (ddr-tdrP + ifcAP) produced 9.67 ± 1.65 µM of equol from DHD, in the presence of NADPH (Figure 4).
{kind=link}