The current investigation determines the influence of severe salt-induced stress and the response of the sensitive (IR29) and moderately tolerant (FL478) rice and how bioinoculation using a multifaceted PGP endophytic bacterium, Methylobacterium oryzae CBMB20, facilitates microbe-mediated salt stress tolerance. Global proteomic differences between the treatments were compared. To establish the impact of salt stress on IR29 and FL478 the non-stressed, non-inoculated rice were compared to the salt-stressed, non-inoculated rice (IR29_SS_NI vs IR29_NS_NI; FL478_SS_NI vs FL478_NS_NI). To assess microbe-mediated salt stress tolerance, the salt-stressed, non-inoculated plants were compared to the salt-stressed, M. oryzae CBMB20 inoculated plants (IR29_SS_IN vs IR29_SS_NI; FL_SS_IN vs FL_SS_NI). Additional information could also be derived from the comparison of FL_SS_NI vs IR_SS_NI and FL_SS_IN vs IR_SS_IN.
The adverse and sometimes detrimental action of salinity in plants is a combinatorial stress of sodium ion (Na+) toxicity, salt-induced osmotic and oxidative stresses complicated by imbalance of ionic homeostasis and deficiency in nutrients (Munns 2002; Evelin et al., 2009; Horie et al., 2012). This combinatorial stress condition individually, cumulatively, or even synergistically induce rice to respond causing dynamic shifts in plant proteomics. Adaptations to salt stress clearly differentiate glycophytes and halophytes allowing halophytes to even thrive in severe stress conditions (Flowers et al., 1977; Hasegawa et al., 2000; Horie et al., 2012), however, understanding responses between tolerant and sensitive genotypes at the molecular level such as the proteomes level potentially allows us to extend the tolerance of important crops, which are generally sensitive to salt stress.
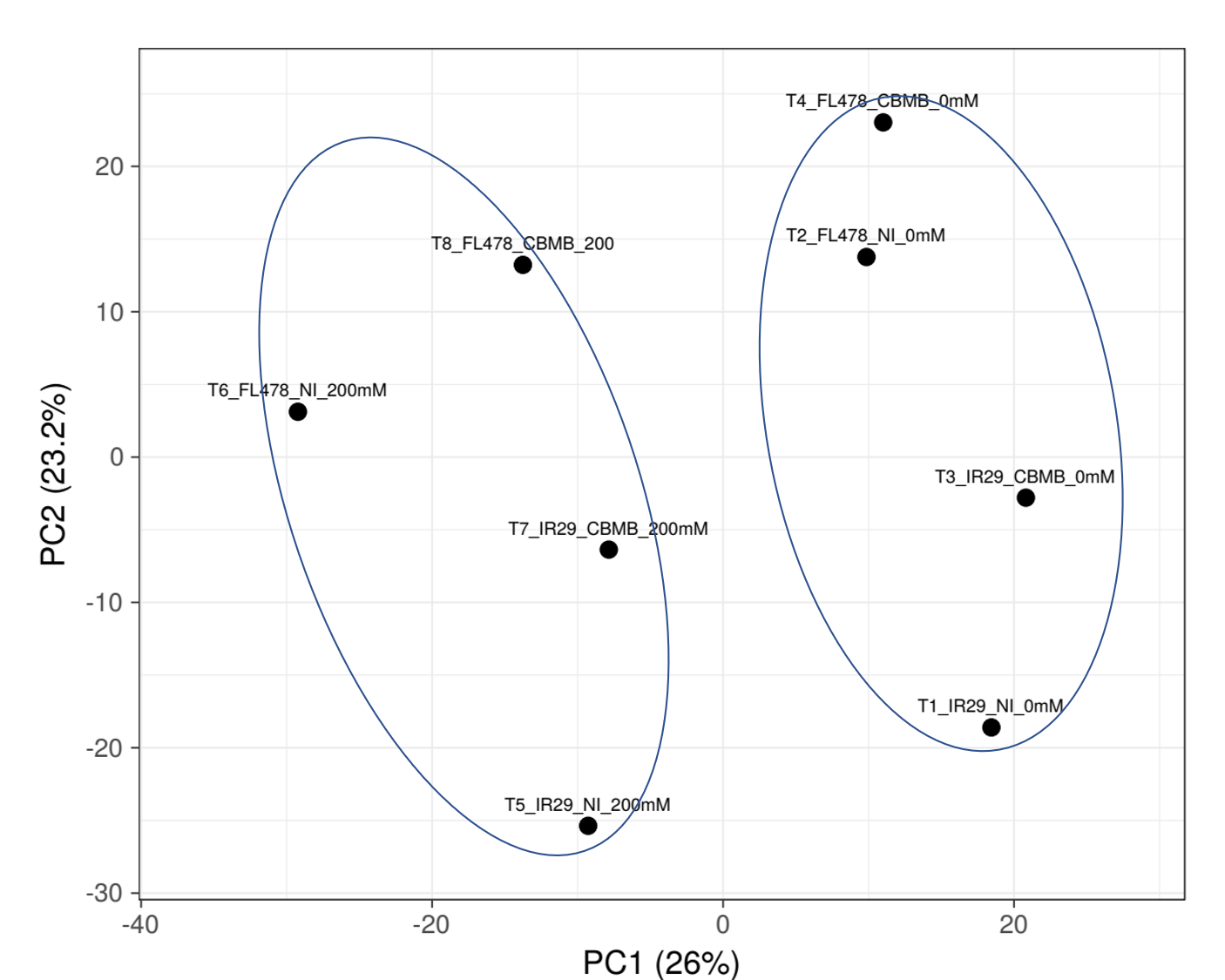
The present study shows that global proteomic differences occur in the tolerant and sensitive rice cultivars under non-stressed and stressed condition and between the inoculated and non-inoculated rice. Looking into the interactions of genotype, salt stress, and bioinoculation on the global proteomes of IR29 and FL478, severe salt stress became a main factor clustering stressed plants and non-stress plants showing an overwhelming proteomic restructuring of rice plants responding to salt stress. The plant genotype becomes the second most important factor affecting global proteomic differences then inoculation as observed in the non-stressed plants. Furthermore, for the salt-stressed plants, it is very interesting that the global proteomes of the inoculated IR29 under salt stress is more similar to the inoculated, salt-stressed FL478 compared to the non-inoculated, salt-stressed FL478. This potentially indicates more significant changes in the global proteomes of the inoculated IR29 under stress comparable to the proteomes of the inoculated, salt-stressed FL478. As was previously reported (Walitang et al., 2023b), M. oryzae CBMB20 bioinoculation on the two rice genotypes under non-stressed conditions have also led to changes in the global proteomes of IR29 and FL478 expanding the proteome’s gene ontology terms observed in both plants with genotype specific proteomic changes especially on the significantly regulated DAPs. In the present study, salt stress and inoculation of plants followed by salt stress imposition have also dramatically altered the proteomes of the two rice genotypes. However, the higher similarity of proteomes of inoculated and stressed IR29 and FL478 indicates a more profound microbe-mediated response in IR29. A similar observation was also seen in a previous investigation where bioinoculation with M. oryzae CBMB20 has led to a greater bacteria-dependent changes in IR29 under stress conditions leading to greater improvement of physiological parameters in the stressed IR29 compared to FL478 (Chatterjee et al., 2019). Another similar observation between inoculated and stressed IR29 and FL478 showing greater responses of the sensitive rice cultivar to bioinoculation compared to the tolerant genotype leading to greater production of osmolytes in IR29, although the inoculant used was Brevibacterium linens RS16, another bacteria that modulates salt stress due to its ACC deaminase production (Ahmed et al., 2021). These observations indicate that inoculation with M. oryzae CBMB20 on the IR29 and FL478 under salt stress conditions lead to microbe-mediated global proteomics changes with a potential more effective microbe-mediated responses on the salt-sensitive rice cultivar.
The changes in global proteomes under stress could be traced to the stress responsive transcriptomics changes. Salt stress induces transcriptional changes to rice cultivars within minutes to hours after imposition of salt stress with potential differences in the gene expression level and even timing responses (Kawasaki et al., 2001). In general, a succession of responses can be observed in salt-stressed plants within a time frame of minutes to weeks following initiation of salt induced stress (Munns 2002; Munns and Tester 2008). The changes in global proteomes between IR29 and FL478 are observed in our study. These differences were also observed in a previous study both in terms of transcriptional and translational level, although the genotypes being compared are IR29 and Pokkali, the original paternal parent of FL478 (Li et al., 2018). Theoretically, FL478 and IR29 share 50% of their genome as IR29 is the original maternal parent of FL478, and the remaining 50% is from the original paternal parent, Pokkali (Walitang et al., 2018, 2019). However, prominent response differences of IR29 and FL478 is evident in a previous study even at the transcriptional level (Walia et al., 2005). The bioinoculation with the endophytic PGPB M. oryzae CBMB20 prior to imposition of salt stress resulted to additional changes and responses in IR29 and FL478. Although transcripts and proteomes do not always correlate due to post transcriptional regulations and changes, the differences in the proteomes of IR29 and FL478 may also be a result of loading efficiency of mRNA onto polysomes (Li et al., 2018) and inherent differences in the timing and efficiency of responses to salt stress (Kawasaki et al., 2001), and other post transcriptional regulation leading to differences in the observed proteomes. The details on the specific DAPs responsive to salt stress and those microbe-mediated responsive DAPs are discussed below.
There are genotype-specific differences and similarities observed when comparing GO terms between IR29 and FL478 in terms of biological processes, cellular compartmentalization and molecular functions. Our results show that under severe salt stress, DAPs related to translation are more abundant in IR29 while those in FL478 are associated with cellular aromatic compound metabolic process and cofactor metabolic process. Other DAPs are also associated with organic acid metabolic process, response to stimulus, and carbohydrate metabolic process observed in both IR29 and FL478. However, if we look into the cellular component in which these DAPs are located, there is an overwhelming abundance of DAPs located in the cytoplasm in IR29 followed by the plastids, membranes and ribosomes in contrast to a more balanced distribution of DAPS in the cytoplasm, plastid and membranes in FL478.
Under salt stress conditions, clearly distinctive differences can be observed between IR29 and the tolerant Pokkali where Pokkali was observed to be more effective in enhancing cell wall integrity, detoxify ROS, translocate molecules and maintain photosynthesis (Li et al., 2018). Theoretically, these observations should also be observed in FL478 as Pokkali is the putative donor of salt tolerance in FL478. Generally, salt stress causes an upregulation of responses (Zhang et al., 2012). However, our observations showed that both sensitive and moderately tolerant genotypes have more downregulated DAPs. This is potentially due to the severity and relatively long duration of severe stress imposition. It is noteworthy to point that FL478 has more significantly upregulated DAPs compared to IR29 indicating its more effective salt stress responses. It is also possible that the presence of more significantly upregulated DAPs in FL478 indicate recovery of downregulated DAPs in the initial response to salinity attaining the same expression or even higher prior to salt stress as observed in its paternal parent, Pokkali (Kawasaki et al., 2001). During the early stages of stress, there is an increased protein synthesis and protein turnover which eventually shifts to induction of stress-responsive transcripts then production of protein with defense-related functions (Kawasaki et al., 2001). The overrepresented DAPs related to translation located in the ribosomes observed in IR29 observed in this study probably indicate the delayed response of the salt-sensitive genotype while transitioning to stress-responsive and defense related functions observed in FL478 with enhanced metabolic related processes.
The GO terms of DAPs in bioinoculated IR29 and FL478 under salt stress condition reflects the overall global proteomic trends observed. In general, the trends in the gene ontology observed in the inoculated, salt-stressed FL478 is also observed in the inoculated, salt-stressed IR29 with slight differences in abundance. These similar patterns were also observed in the cellular localization, molecular functional roles and biological processes of the gene ontology terms of DAPs.
Proteome responses to severe salt stress in IR29 and FL478 rice genotypes: salt stress-responsive proteins
Severe and long-term salt stress adversely affects similar mechanisms in plants, whether they are sensitive or tolerant, and the differences in responses determines their stress tolerance in contrast to clear adaptations of halophytes (Flowers et al., 1977). Key features in the appearance of salt tolerance are the more efficient and effective management of stress responses rather than creation of adaptations uniquely seen only in specialized plants such as halophytes (Kawasaki et al., 2001; Munns and Tester 2008). Severe salt stress in our study have shown specific salt stress-responsive proteins common to both IR29 and FL478 indicating similar features and processes influenced by salt stress in terms proteins. We have detected five DAPs significantly upregulated in both IR29 and FL478. These include the Usp domain-containing protein (OsI_02150), glutaredoxin domain-containing protein (OsI_37657), aldehyde dehydrogenase (NAD(+)) (OsI_31526) and peptidyl-prolyl cis-trans isomerase (OsI_00098) as well as an uncharacterized protein (A2X9N1). The universal stress protein (Usp) domain containing protein (OsI_02150) has a fold change (FC) upregulation of 3.93 in IR29 and 4.82 in FL478 indicating its clear importance in salt stress response in both genotypes. The glutaredoxin domain-containing protein (OsI_37657), aldehyde dehydrogenase (NAD(+)) (OsI_31526) and peptidyl-prolyl cis-trans isomerase (OsI_00098) also have relatively higher FC values. It is also noteworthy to point that the uncharacterized protein A2X9N1 significantly upregulated in FL478 has an FC value of 7.45, the most upregulated proteins detected in IR29 and FL478 induced by salt stress. Furthermore, the greater FC values in FL478 compared to the IR29 of these stress-related proteins such as in the case of Usp domain containing protein, aldehyde dehydrogenase, peptidyl-prolyl cis-trans isomerase and the uncharacterized protein A2X9N1 supports the notion that the moderately salt tolerant FL478 is more efficient and more responsive in their regulation of salt stress leading to higher tolerance. These are in line with previous studies comparing IR29 and FL478 (Walia et al., 2005; Cotsaftis et al., 2011, Senadheera et al., 2012, Mirdar Mansuri et al., 2019), IR29 and Pokkali (Kawasaki et al., 2001; Cotsaftis et al., 2011; Li et al., 2018), or other cultivars (Kumari et al., 2009) associated to salt stress defense and related DAPs. The universal stress protein (Usp) is a multi-stress responsive protein conserved and providing stress resistance to relatively diverse organisms (Bhuria et al., 2019; Chi et al., 2019). The aldehyde dehydrogenase is active in detoxification, antioxidant and regulatory functions (Shortall et al., 2021), but it catalyzes dehydrogenation of D-glyceraldehyde 3-phosphate into 3-phospho-D-glycerate and NADH in rice. The aldehyde dehydrogenases are also involved in the oxidation of aldehydes to carboxylic acids in rice (Gao and Han 2008). The detected glutaredoxin domain-domain containing protein detected in both IR29 and FL478 is involved in reducing GSH-thiol disulfides in rice associated with ion binding and transport. They are involved in the oxidative response in many plants directly reducing peroxides, dehydroascorbate, peroxidoxins and protecting thiol groups (Rouhier et al., 2004, Rouhier et al., 2006). The peptidyl-prolyl cis-trans isomerase detected in the present study can accelerate folding of proteins. Two variants were detected in FL478 and the one located in the chloroplast is also associated to lateral root morphogenesis, photosystem II assembly and photosystem II stabilization. In higher plants, they assist in protein folding and those found in chloroplasts, mitochondria, and cytosol are light-controlled and are stimulated by heat, cold and high salt concentration (Blecher et al., 1996) while they participate in extracellular matrix development on the cell surfaces (Lin et al., 2019). The uncharacterized protein (A2X9N1) is related to ATP synthesis and ion transport. These proteins are potentially critical in both IR29 and FL478 responding to severe salt stress conditions.
The common downregulated DAPs in both IR29 and FL478 are associated to diverse biological processes including photosynthesis (NADPH-protochlorophyllide oxidoreductase (OsI_17934), Clu domain-containing protein), transcription related and chromosomal stability (histone H4), carbohydrate metabolism (sucrose synthase (OsI_11950), UDP-glucose 6-dehydrogenase (OsI_13603)), molecular chaperones (CCT-eta (OsI_24243) and metabolism (adenylosuccinate synthetase, chloroplastic (PURA), UDP-glucose 6-dehydrogenase (OsI_13603). In some previous studies, there were few detected common downregulated transcripts between IR29 and FL478 (Walia et al., 2005) and IR29 and Pokkali (Li et al., 2018). In this study, we have detected more commonly downregulated proteomes in IR29 and FL478 potentially due to their common response to severe salt stress condition. In addition, our result showing downregulation of many DAPs in both IR29 and FL478 potentially indicates harmful and possibly detrimental effect of severe salt stress on both genotypes.
Aside from the common significant DAPs between IR29 and FL478, genotype-specific significantly regulated DAPs also provide information on the proteomic responses of rice and the potential mechanisms of salt stress through proteomes. Most of the significantly upregulated DAPs in FL478 can be broadly categorized into four clusters particularly those related to carbohydrate and energy metabolism, photosynthesis, stress and defense, and metabolism. While in IR29, protein synthesis proteins are abundant followed by stress and defense-related proteins, metabolism of carbohydrates and photosynthesis. In addition, most of the downregulated DAPs are also associated to the synthesis of protein, transcription, metabolism, metabolism of carbohydrate and photosynthesis. It can be noted that the biological processes in which most of the upregulated DAPs are associated with are also the same with those that were downregulated particularly protein synthesis, carbohydrate metabolism and photosynthesis. This indicates that one of the responses to salt tolerance in IR29 seems to balance the processes greatly depreciated by salt stress by upregulating similar functional proteins that were downregulated by salt stress.
Aside from the stress and defense related proteins discussed above commonly upregulated in both IR29 and FL478, there are greater representation of FL478 genotype-specific proteomes associated to photosynthesis, carbohydrate metabolism and stress and defense. Photosynthesis is severely influenced by salt stress further developing to critically detrimental effects in plants (Hasegawa et al., 2000). Photosynthesis is linearly affected by increasing sodium concentration in the leaves particularly in older plants which concurrently affect stomatal aperture and gas exchange as well as carbon dioxide fixation (Yeo et al., 1985; Horie et al., 2012). Osmotic effects of salt in the rhizosphere can also severely impact photosynthesis in plants (Tattini et al., 1995; Munns 2002). Looking at the specific proteins in FL478, there are more DAPs upregulated compared to a few DAPs in IR29 related to photosynthesis. These overrepresented photosynthesis-related DAPs are associated to thylakoid and chloroplast integrity and photosystem II assembly and stabilization. Salt-tolerant rice cultivars are known to more effectively manage photosynthesis-related aspects compared to their salt-sensitive counterparts (Li et al., 2018) probably as a result of their higher chlorophyll pigment contents (Walia et al., 2005) leading to photosynthesis recovery under prolonged stress in contrast to the sensitive rice genotype (Walia et al., 2005). These studies implicate that there is less severe stress in FL478 as observed in Pokkali (Li et al., 2018)
Inoculation with Methylobacterium oryzae CBMB20 restored the abundances and potential functions of significantly downregulated proteomes under severe salt stress conditions
The application of M. oryzae CBMB20 profoundly changed the proteomes of IR29 and FL478 even under ideal growing environment (Walitang et al., 2023b). One of the most prominent changes observed after inoculating IR29 and FL479 followed by salt stress imposition is the ‘restoration’ of function of proteins. We have observed common and genotype-specific DAPs that were upregulated in salt-stressed rice plants applied with M. oryzae CBMB20 while their counterparts were severely downregulated the salt-stressed, non-inoculated rice plants.
Both IR29 and FL478 showed three major DAPs with significant upregulation when they were applied with M. oryzae CBMB20 then subsequently exposed to salt stress. Like our results, sucrose synthase and peroxidase enzymes were downregulated under salt stress conditions in IR29 and Pokkali (Kawasaki et al., 2001) and in IR29 (Li et al., 2018). Sucrose synthase is also downregulated due to salt-induced stress (Peng et al., 2009; Pang et al., 2010; Yu et al., 2011). The peroxidase enzyme has unpredictable regulation in different plants under stress as it can be up- or down-regulated. The third DAP, NADPH-protochlorophyllide oxidoreductase (OsI_17934) was observed to be downregulated in Arabidopsis thaliana leaf microsomal membrane due to salt-induced stress (Pang et al., 2010). However, inoculation with M. oryzae CBMB20 restored the functions of these proteins as their abundance and fold change increased in both IR29 and FL478.
The DAP NADPH-protochlorophyllide oxidoreductase (OsI_17934) is involved in the phototransformation of protochlorophyllide to chlorophyllide, an essential process in the transformation of chloroplast from etioplasts (Apel et al., 1980). This enzyme was also observed to be upregulated in non-stressed, M. oryzae CBMB20 inoculated IR29 (Walitang et al., 2023b). Our current result showed very high upregulation of this DAP due to M. oryzae CBMB20 in salt-stressed rice plants. Plant pigments (chlorophyll a, b, and carotenoids) were significantly increased under normal and under salt stress in a japonica cultivar inoculated with M. oryzae CBMB20 (Roy Choudhury et al., 2022). Photosynthetic rate and PSII efficiency in IR29 and FL478 also improved due to M. oryzae CBMB20 (Chatterjee et al., 2019). These improvements in the photosynthetic traits under salt stress when plants were applied with M. oryzae CBMB20 could be due to the upregulation and restoration of DAPs associated with the chloroplast or photosynthesis as in the case of NADPH-protochlorophyllide oxidoreductase.
Sucrose synthase (OsI_11950) cleaves sucrose to UDP-glucose and fructose. The end products of sucrose breakdown by sucrose synthase becomes available to diverse metabolic pathways generation of energy, production of primary metabolites and complete carbohydrate biosynthesis (Stein and Granot, 2019). In rice, it is involved in some housekeeping roles, cellulose synthesis, carbon allocation, and stress response (Hirose et al., 2008). The upregulation of two variants of sucrose synthase in both IR29 and FL478, Sucrose synthase (OsI_11950) and Sucrose synthase (OsI_22003), indicates a potential improvement or restoration of functions attributed mainly to carbohydrate metabolism and energy production. In a previous study in salt stressed Oryza sativa ssp. japonica inoculated with M. oryzae CBMB20, majority of DAPs belong to carbohydrate metabolic process (Roy Choudhurry et al., 2022). A similarly observation can also be seen in this study.
The peroxidase upregulated in IR29 in mainly involved in the catabolism of hydrogen peroxide and oxidative stress response while the two peroxide variants observed in FL478 are also involved in lignin degradation and synthesis, catabolism of auxin, and response to stresses including invasion of pathogen, oxidative damage and physical damage. Upregulation of peroxidase in Pokkali was mainly observed during the stress compensation period where upregulation of defense against reactive oxygen species predominates (Kawasaki et al., 2001). In addition, the peroxidase enzyme observed in FL478 potentially contributes to lignin biosynthesis which could promote additional cell wall rigidity, similar to the observation in Pokkali (Li et al., 2018). The observed M. oryzae CBMB20-induced upregulation of antioxidant activities protecting plants against salt stress was also seen in previous studies (Chanratana et al., 2019). Supporting the functions of the peroxidase enzyme, the genotype-specific enzyme xyloglucan endotransglucosylase/hydrolase (OsI_28350) upregulated in the M. oryzae CBMB20 inoculated FL478 also contributes to cell wall reconstruction indicating that inoculation with M. oryzae CBMB20 under salt stress enhances cell wall reconstruction adding another layer of defense response to salt-induced stress. Pokkali can sustain better plasticity of the cell wall under salt stress compared to IR29 (Li et al., 2018). However, it seems that inoculation with M. oryzae CBMB20 have enhanced this vital response in FL478 in the current study.
The other genotype-specific DAPs whose abundances were significantly upregulated potentially restoring their activities are associated to different functions. In IR29, two DAPs are associated to protein synthesis: the ribosomal component 40S protein and the eukaryotic translation initiation factor, and another DAP is associated to purine nucleotide biosynthesis (adenylosuccinate synthetase). On the other hand, the genotype-specific DAPs with restored function in M. oryzae CBMB20 inoculated FL478 are involved with salt stress tolerance (fasciclin-like arabinogalactan protein), protein synthesis (NAC-A/B domain containing protein), and one carbon metabolism (methylenetetrahydrofolate reductase). The restoration of abundances of downregulated proteins due to the application of M. oryzae CBMB20 together with the other genotype-specific significantly upregulated DAPs indicates the essential impact of inoculation in rice similarly observed even under non-stress conditions (Walitang et al., 2023b) and in Oryza sativa ssp. japonica under severe salt stress (Roy Choudhury et al., 2022).
Hypothetical mechanisms of action leading to the dynamic proteomic responses of the M. oryzae CBMB20 inoculated rice plants under severe salt stress
Methylobacterium oryzae CBMB20 is a multifunctional PGPB with growth promoting abilities including N2 fixation, phytohormone production (IAA, cytokinin) (Lee et al., 2006; Ryu et al., 2006), 1-aminocyclopropane-1-carboxylate (ACC) deaminase production (Madhaiyan et al., 2007), and thiosulfate oxidation (Anandham et al., 2007). Under salt stress conditions, there are potential mechanisms of action by this multifaceted PGPB.
M. oryzae CBMB20 can enhance health of stressed plants including pathogen attack (Indiragandhi et al., 2008; Yim et al., 2013, 2014), heavy metal stress (Madhaiyan et al., 2010), and salt stress (Chanratana et al., 2018; Chatterjee et al., 2019; Roy Choudhury et al., 2022; Roy Choudhury et al., 2023a, b). This amazing feat by Methylobacterium oryzae CBMB20 is mainly due to its ACC deaminase activity under stress conditions. Methylobacterium oryzae CBMB20 is capable of regulating stress ethylene by sequestering and hydrolyzing the ethylene precursor, ACC, to α-ketobutyrate and ammonia through the action of its ACC deaminase (Madhaiyan et al., 2007). In our current investigation, the restoration of abundances of numerous downregulated DAPs under severe salt stress in both IR29 and FL478 after application of M. oryzae CBMB20 is potentially due to lowering of stress via ethylene reduction. This potentially allowed both IR29 and FL478 to recover and upregulate their DAPs initially downregulated due to salt stress.
Aside from the action of ACC deaminase, it is also still possible that M. oryzae CBMB20 can maintain plant growth promotion through the direct mechanisms mentioned above even under stress conditions.
Another potential mechanism as recently supported by study (Roy Choudhury et al., 2023b) is the involvement of ethylene-induced pathogenesis related signaling. Priming may also be involved in this case. In priming, the sensitization of the whole plant for stronger defense responses, mainly characterized by faster, more robust and more efficient activation of cellular defenses is well established in PGPB inoculated plants subsequently challenged by pathogens or attacked by herbivores (Pieterse et al., 2014). The enhanced protection of M. oryzae CBMB20 on pathogen-challenged plants were already observed particularly against R. solanacearum (Yim et al., 2013), Xanthomonas campestris (Yim et al., 2014)d syringae pv. tomato (Indiragandhi et al., 2008), although all these studies have mainly attributed this to ACC deaminase and the regulation of stress ethylene. In a recent study, stress and defense responses have been activated and modulated by the application of M. oryzae CBMB20 under normal conditions in both IR29 and FL478 when M. oryzae CBMB20 endophytically colonized the rice plants (Walitang et al., 2023b). We are proposing that stress and defense responses have been primed due to the application of the M. oryzae CBMB20 inducing systemic resistance leading to a more effective response when the rice cultivars were subsequently exposed to severe salt stress leading to the observed dynamic proteomic responses observed in this study.
{kind=link}