1. PEDV infection affects changes in the intestinal flora and ILC3 in piglets.
In our previous work, we found a large number of immune cells in the piglet intestine [20], and through flow cytometric sorting and single-cell sequencing (scRNA-seq), we found that in Lin− cells, there were a large number of ILCs cells (Fig. 1A), in which ILC3 expressing the signature transcription factor RORC was absolutely dominant (Fig. 1B), and also expressing, IL7R, AhR and other surface molecules, and secreted corresponding cytokines such as IL22 and IL17. The expression of these genes is important for the regulation and protection of intestinal homeostasis (Fig. 1C).
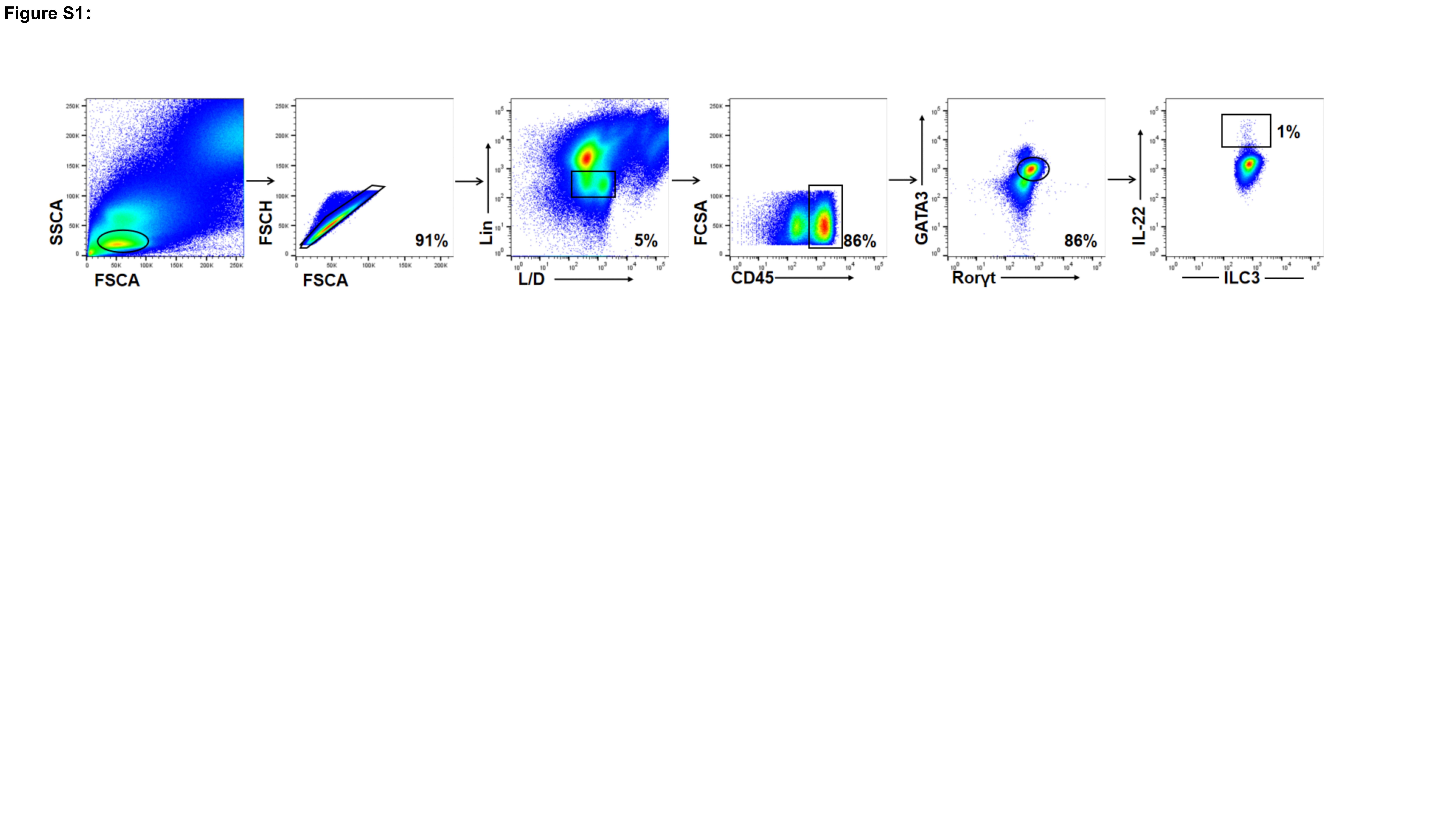
Through scRNA-seq analysis, we found that PEDV infection in piglets led to a significant decrease in the number of immune cells in the jejunum, including T cells and ILC3 (Fig. 1D, F, G). In contrast, non-immune cells, such as epithelial cells, exhibited abundant proliferation during the infection. (Fig. 1D, H). Despite the significant decrease in the number of ILC3 cells in the jejunum, the secretion of IL-22 was markedly increased (Fig. 1E, I). These results suggest that the increased secretion of IL-22 by ILC3 cells may play a crucial role in conferring resistance against PEDV during PEDV infection. Our flow cytometry assay results were consistent with the single-cell results, the flow gating strategy in Fig. S1A. These results also confirmed a significant decrease in the number of ILC3 cells in the intestinal lamina propria of PEDV-infected piglets, accompanied by an up-regulation of IL-22 secretion by ILC3 cells (Fig. 1J). Furthermore, our findings revealed significant alterations in the flora of the jejunum in PEDV-infected piglets. At both the order level and the genus level, there was a notable decrease in the levels of Lactobacillales orders and Lactobacillus genus (Fig. 1K, L).
To further investigate the underlying cause of the significant changes in ILC3 during PEDV infection, we aimed to discern whether these changes were primarily driven by viral infection or influenced by alterations in the bacterial flora. To explore this, we conducted an experiment involving antibiotic treatment to manipulate the structure of the flora. By observing the effects of this flora change on the body's immunity, we aimed to gain insights into the intricate relationship between the microbial composition and the immune response.
2. Alteration of intestinal flora may affect ILC3 development.
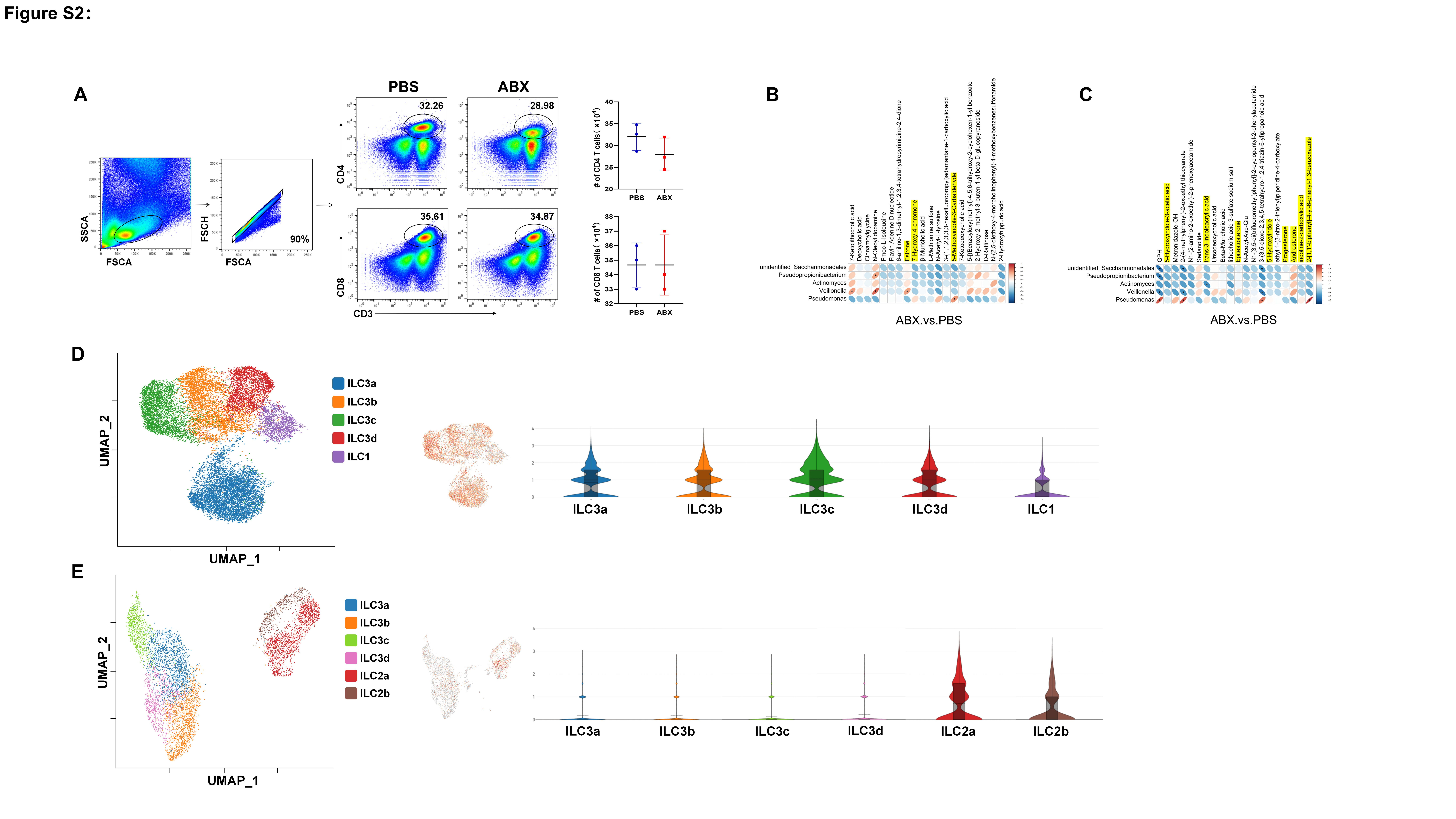
In our study, we initially established a dysregulation model of piglets' microflora. We observed significant impacts on the immune development of piglets with dysflora. Specifically, the number of CD4-T cells in the jejunum lamina propria of the jejunum decreased (Fig. S2A). Additionally, the development of ILC3 was even more severely affected, with a significant decrease in cell numbers and an increase in IL-22 secretion (Fig. 2A). Meanwhile, antibiotic treatment resulted in the decrease of flora richness and lactobacillus number in the jejunum of piglets (Fig. 2B, C). Linear discriminant analysis Effect Size (LEfSe) analysis identified significantly different taxa between groups, which were significantly enriched from the point of view of order, genus, and species, respectively, belonging to Lactobacillales, Lactobacillaceae, and Lactobacillus (Fig. 2D, E, F, G).
The above results suggest that the number of immune cells, especially ILC3, was significantly affected under both PEDV infection and antibiotic treatment conditions and that this change persisted when only the flora was changed. The flora may play a very important role in the interactions with ILC3, and if there is a regulatory effect of the flora on ILC3, by what pathway does the flora regulate ILC3? Up-regulation of indole analogs, including bisindolylmaleimide I and indoxyl sulfate, has been reported in PEDV infection [3]. Non-targeted metabolomics was further analyzed for the metabolites in jejunum samples. As a result, we found significant changes in some AhR-related regulatory substances in both positive and negative ion modes (Fig. 2H, I, J, K), especially many indoles that have been repeatedly shown to have a significant activating effect on the AhR. We further performed enrichment analyses and found a significant enrichment in important immune-regulatory pathways such as tryptophan. To investigate whether differential microbial communities produce differential metabolites, we conducted a combined analysis of 16S and metabolomics. The results revealed a significant decrease in the abundance of regulatory substances secreted by probiotic bacteria, leading to a reduction in their overall levels. However, unfortunately, we did not directly associate the metabolites with the production by Lactobacillus (Figure S2B, C).
So far, the expression of the AhR receptor has been studied in immune cells in humans and mice, but its expression has not been reported in pigs. Furthermore, the expression of AhR in immune cells in the intestine is unknown. We found that immune cells in the porcine intestine highly expressed AhR receptors by single-cell analysis (Fig. 2L), and further analysis revealed that ILC3 in the intestine expressed high levels of AhR (Fig. 2M). In human intestinal ILC, both ILC3 and ILC1 highly expressed AhR receptors (Fig. S2D), whereas only ILC2 expressed AhR in the murine upper intestine (Fig. S2E). These results suggest that porcine and human intestinal ILC3 exhibit high levels of AhR activation, indicating that ILC3 in the intestine may respond significantly to stimulation by AhR ligand-related substances.
3. Feeding LGG can enhance the piglets' immunity against PEDV infection.
To explore whether Lactobacillus can resist PEDV infection in the gut, we selected the standard Lactobacillus strain LGG to supplement piglets with an intestinal flora imbalance and then carried out a PEDV challenge experiment. Our objective was to determine whether feeding LGG could enhance intestinal immunity, specifically by activating ILC3 cells, thereby promoting intestinal homeostasis and conferring resistance against PEDV infection.
Our results found that LGG supplementation significantly reduced the pathological changes of PEDV infection in piglets, and the intestinal tract was healthy in the CON group. The villi of the jejunum of piglets in the ABX + LGG + PEDV group were shortened and atrophied, and the epithelial cells were mildly diseased. The villi of the jejunum in piglets from the ABX + PEDV group exhibited shortened, fragmented, and broken structures. Additionally, the intestinal epithelial cells underwent typical histopathological changes such as vacuolization, fusion lesions, necrosis, and detachment following PEDV infection (Fig. 3A). The ratio of villus height to crypt depth (VH/CD) of the jejunum villi of piglets in each infection group was found to be significantly lower in the ABX + PEDV group than in the ABX + LGG + PEDV group (Fig. 3B). The diarrhea scoring results proved that feeding LGG alleviated diarrhea in piglets (Fig. 3C). Next, we found significant differences in viral loads in the duodenum, jejunum and ileum between the ABX + LGG + PEDV group and the ABX + PEDV group, especially in the jejunum and ileum, where the viral copy number was significantly decreased after feeding LGG (Fig. 3E).
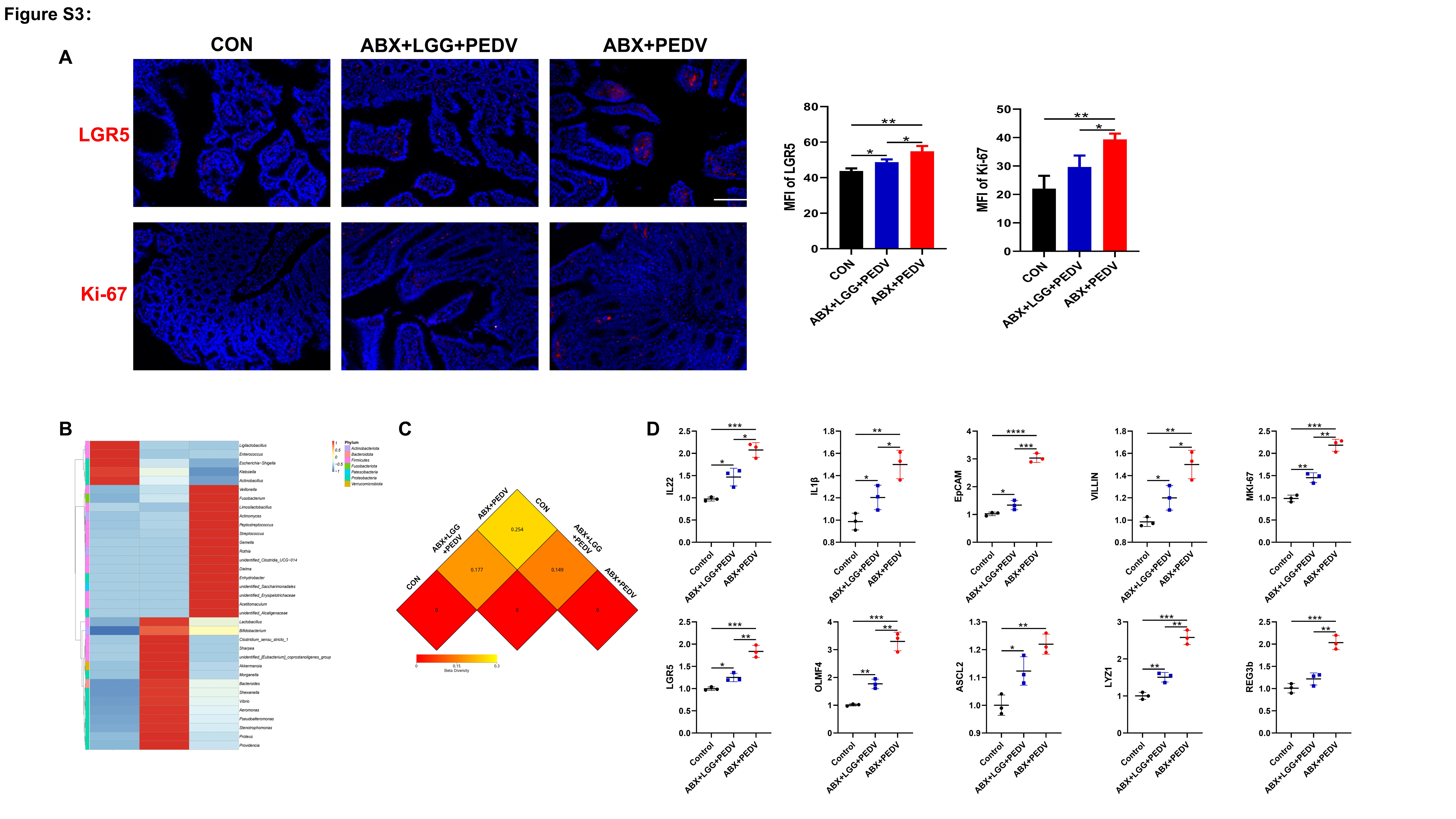
Flow results also demonstrated a significant increase in the amount of ILC3 in the intestines of the ABX + LGG + PEDV group, which was close to that of the CON group and significantly higher than that of the ABX + PEDV group. The secretion of IL-22 was also significantly lower than that of the ABX + PEDV group, suggesting that the inflammatory state of the jejunum intestinal tract of the piglets was ameliorated in response to the infection (Fig. 3D). A dramatic improvement also occurred in the abnormal activation state of the intestinal tract through stimulation by feeding LGG to promote proliferation of epithelial cells and the production of IL-22 after viral infection, including the expression of proteins such as EpCAM and VILLIN in the intestinal tract (Fig. 3F). There was also a significant reduction in the cell proliferation-related proteins LGR5 and Ki-67 (Fig. S3A).
Feeding LGG also significantly increased the abundance of intestinal flora in piglets, particularly the number of the Lactobacillus genus, and elevated the content of other probiotics, such as Streptococcus spp (Fig. 3G). The alpha diversity index of the microbial community shows a significant increase (Fig. 3H). The heatmap analysis of the microbial community at the genus level and the results of beta diversity analysis also suggesting that feeding LGG can promote the recovery of the intestinal flora (Figure S3B; C)..
Our results at the mRNA level were consistent with the results at the protein level, revealing that inflammation-related genes such as IL-22 and IL-1β, the epithelial gene EpCAM, the villi gene VILLIN, the proliferation and differentiation-related gene MKI-67, the ISC labeling genes Lgr5, Ascl2, Olfm4, and the Paneth cell activation genes Lyz1 and REG3b were all significantly reduced (Figure S3D).
4. EVs ( LGG) promote ILC3 cell activation and secretion of IL-22.
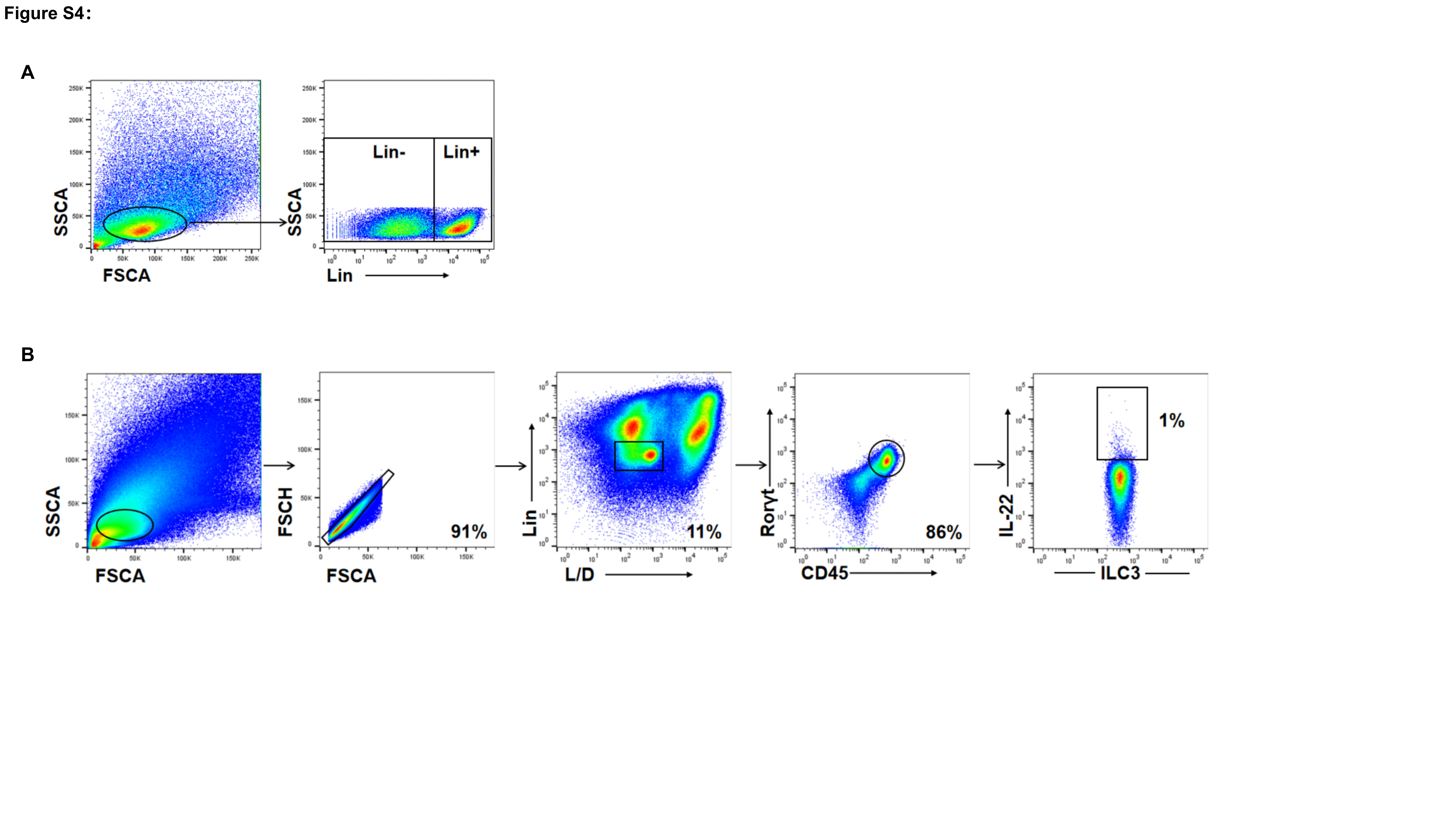
We first successfully constructed an in vitro culture model of ILC3 (Fig. 4A). The ILCs were collected by magnetic bead sorting (Fig. S4A) and performed a flow cytometry assay under different stimulant-inducing conditions, and developed a gating strategy (Fig. S4B). We found that LGG-secreted extracellular vesicles (EVs) could significantly promote ILC3 proliferation and secretion of IL-22 (Fig. 4B).
We enriched and collected the extracellular vesicles (EVs) through ultrafast centrifugation and found the diameter of the EVs to be around 70 nm by nanoflow cytometry (Fig. 4C). The signature horseshoe-shaped structure of the EVs, which contained a large amount of LGG metabolites, was visualized under an electron microscope (Fig. 4D). Subsequently, we conducted untargeted metabolome detection. In total, 326 substances were identified in the positive mode (NEG), while 414 substances were detected in the negative mode (POS). We observed a significant presence of lipids and lipid-like molecules, as well as organic acids and derivatives, in both the POS and NEG modes (Fig. 4E). We found a wide range of substances that can interact with AhR, including indole compounds such as L-5-Hydroxytryptophan and Indole-3-acrylic acid, as well as several ketone compounds, and some ketones. Research has indicated that, apart from indoles, ketones can also interact with AhR [21]. We conducted KEGG enrichment analysis on the total metabolites and found significant enrichment in the Metabolism pathway. Additionally, enrichment was observed in pathways such as Cellular Processes, Environmental Information Processing, and Genetic Information Processing (Fig. 4F).
5. EVs ( LGG) promote ILC3 cell activation to resist PEDV through AhR receptor
We found that both LGG-secreted extracellular vesicles (EVs) and Indole-3-carbinol (I3C) could significantly promote ILC3 proliferation and secretion of IL-22 through activation of the AhR. The effect disappeared when the AhR antagonist CH-223191 (CH) was added (Fig. 5A).
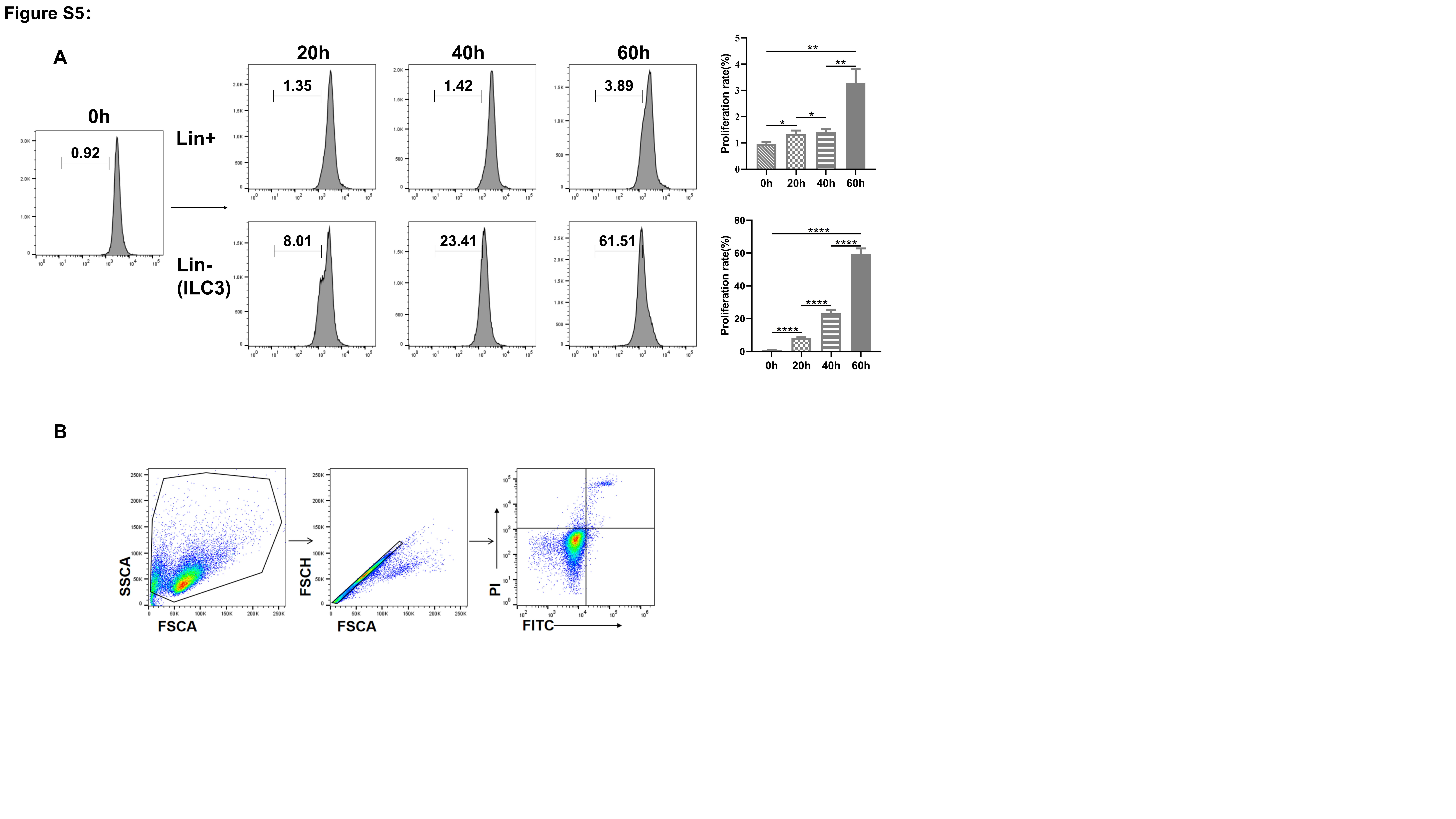
Previous correlational studies have indicated the possibility of ILC transformation within the organism, particularly when induced by specific conditions [22]. To investigate whether the change in the number of ILC3 is due to the transformation of other immune cells, we conducted magnetic bead sorting on ILCs cells and subsequently examined them using SFSE staining. We discovered that ILC3 is the primary cell population within ILCs where proliferation occurs. The proliferation of ILCs cells dominated by ILC3 was significantly stronger than that of other non-ILC cells (Fig. S5A). The expression of ILC3-related cytokines, including IL-22, IL-17A, IL-17B, CXCL2, and CXCL8, were all found to be significantly up-regulated (Fig. 5B/C).
Our study investigated whether the activation of ILC3 and secretion of IL-22 promoted by EVs are effective in enhancing the resistance of porcine epithelial cells to PEDV infection. We established a co-culture model of ILC3 and IPEC-J2 cells (Fig. 5D) and observed that IL-22 secreted by EVs-promoted ILC3 had a positive impact on IPEC-J2 cell proliferation and the activation of STAT3 (Fig. 5E). Furthermore, when EVs and ILC3 were added to co-cultured IPEC-J2 cells after PEDV infection, it significantly influenced the outcome of PEDV infection by preventing apoptosis of IPEC-J2 cells (Fig. 5F), the gating strategy in Fig. S5B.
These results suggest that EVs derived from LGG can stimulate the proliferation and secretion of IL-22 by ILC3 through the AhR receptor on ILC3. This process promotes the proliferation and activation of epithelial cells and enhances the expression of the STAT3 gene. Therefore, it alleviates IPEC-J2 apoptosis caused by PEDV infection.
6. EVs ( LGG ) can promote the development of porcine intestinal organoids through activating ILC3s to secrete IL-22.
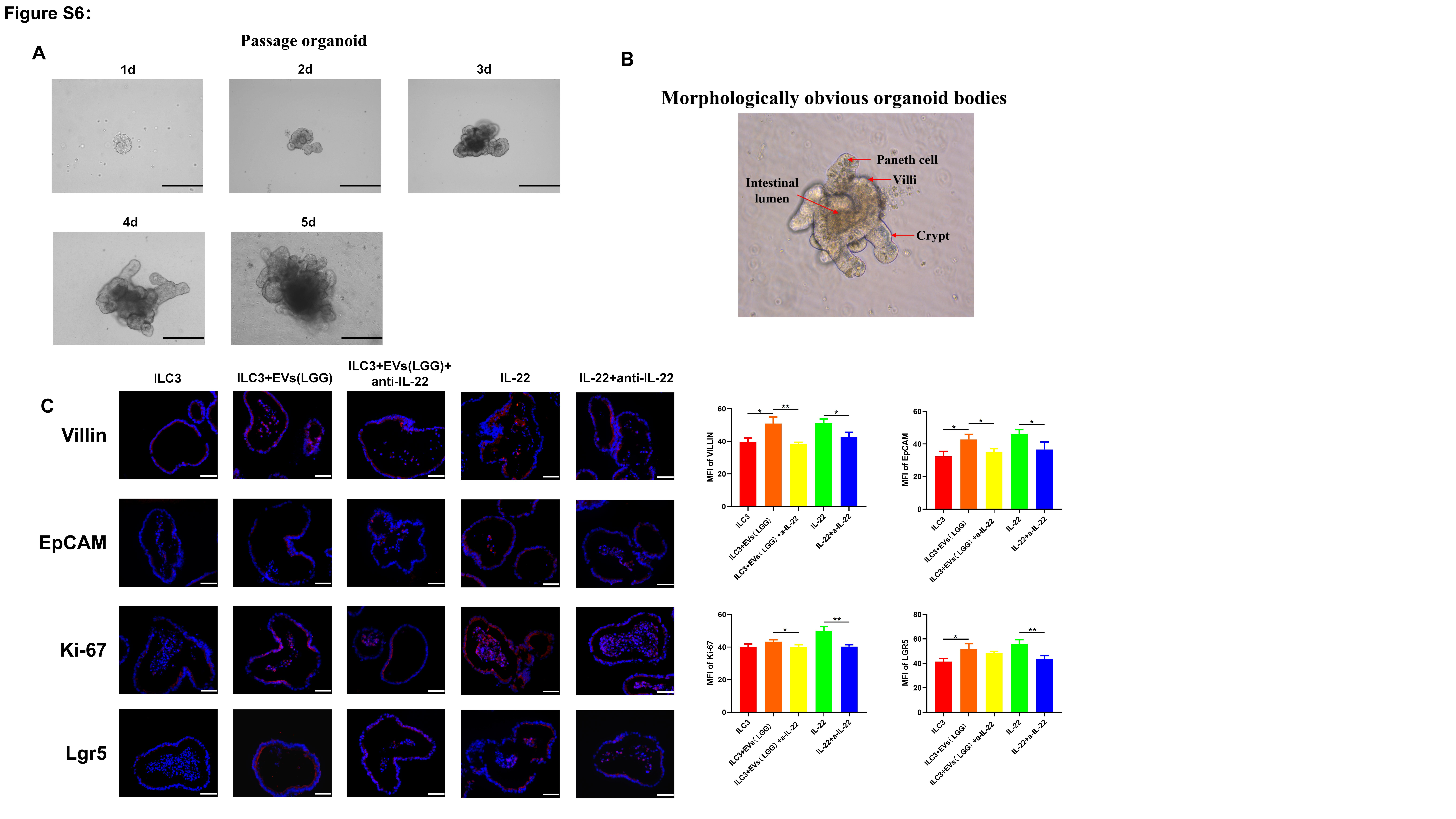
We successfully constructed an in vitro model of organoids from the jejunum of piglets. The size of isolated intestinal crypts was about 15 µm, and they were cultured with matrigel. The first generation of organoids grew slowly and developed into mature bodies with a diameter of about 200 µm by the eighth day and were continued to be cultured. Then they died and fragmented, and displayed a large number of buds in 5–8 d and could be propagated through passaging culture(Fig. 6A). Subsequently, the cultured organoids grew rapidly after passaging, reaching a size of 100 µm in three d and matured into bodies with a diameter of 200 µm within 5 d (Fig. S6A). Similar to the structure of human and murine intestinal organoids, porcine intestinal organoids also consisted of Intestinal Iumen, Villi, Crypt, Paneth cell and other parts (Fig. S6B).
Previous experiments have shown that EVs can stimulate the secretion of IL-22 from ILC3. Therefore, it is worth investigating if IL-22, secreted by ILC3, can promote the growth of organoids. In our study, we initially supplemented the intestinal organoid medium with HK LGG and EVs derived from LGG (Fig. 6B). We then monitored the number of organoids, organoid size, and germination rate throughout the organoid growth process. Interestingly, we observed that the addition of HK LGG and EVs alone did not have any noticeable effect on the development of organoids (Fig. 6D).
In the next phase of our study, we successfully established a co-culture system involving intestinal organoids, stimulatory molecules, and ILC3 cells (Fig. 6C). Initially, we attempted to culture ILC3 cells by adding them directly to the matrigel. However, this method proved unsuitable for the survival of ILC3 cells and resulted in severe cell death and fragmentation within 24 h. To overcome this issue, we performed co-cultures using a transwell system with a pore size of 0.4 µm. We renewed the upper layer of ILC3 cells every 24 h. Notably, we observed that the growth of the organoids was significantly enhanced in the co-culture system when EVs + ILC3 and IL-22 were added. Within the first two d, the organoids experienced rapid growth, reaching a size of 70 µm. Subsequently, they started to exhibit significant budding and divided into multiple crypts on the third day. The organoids continued to grow rapidly, whereas the promotional effect of EVs and IL-22 on organoid growth disappeared upon the addition of an antibody specific to IL-22. Importantly, the number of organoids was not affected (Fig. 6E). Comparing the experimental group (EVs + ILC3, IL-22) with the control group (including ILC3, ILC3 + EVs(LGG) + anti-IL-22, IL-22 + anti-IL-22), we found that the experimental group significantly promoted the growth of the organoids. Notably, a significant difference was observed on the third day during germination, and by the sixth day, the size of the organoids in the experimental group showed significant differences compared to the control (Fig. 6F).
Next, immunofluorescence results revealed significant expression of proliferation-related proteins in organoids after addition of co-cultures of EVs + ILC3 and IL-22, including increased expression of 5-Ethynyl-2′-deoxyuridine (EdU) (Fig. 6G), the ISCs activation-related protein LGR5, and the cell proliferation-related protein Ki67; a significant increase in expression of the epithelial Significantly increased expression of the epithelial protein EpCAM and the chorionic protein VILLIN was also found (Fig. S6C). qPCR results also identified the epithelial gene EpCAM, the chorionic gene VILLIN, the proliferation and differentiation-associated gene MKI-67, and the ISCs marker genes Lgr5, Ascl2, and Olfm4 in the class of organoids following the incorporation of co-cultures of EVs + ILC3 and IL-22; and Paneth cell activation genes Lyz1 and REG3b were significantly increased (Fig. 6H). These results demonstrated that the EVs of LGG could significantly promote the growth and development of organoids and activate ISCs, Paneth cells, epithelial cells, and other cells in organoids.
Our study found the metabolites of LGG are rich in AhR ligands, such as indole compounds. And provides insights into the interaction between ILC3s in the porcine small intestine and Lactobacillus, aiming to understand better how they interact. We have demonstrated that oral LGG is a potential approach for preventing PEDV infection in pigs. It can alleviate intestinal inflammation, intestinal damage, and clinical diarrhea symptoms caused by PEDV infection. We found that EVs (LGG) can activate ILC3s and promote IL-22 secretion, thereby influencing intestinal stem cell regeneration and epithelial protection. Ultimately, this helps to purify the gut environment and resist PEDV infection. In summary, our research demonstrates that oral LGG promises to be a potential method for preventing PEDV infection in pigs. Furthermore, it opens up new avenues for preventing PEDV infection in piglets.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}