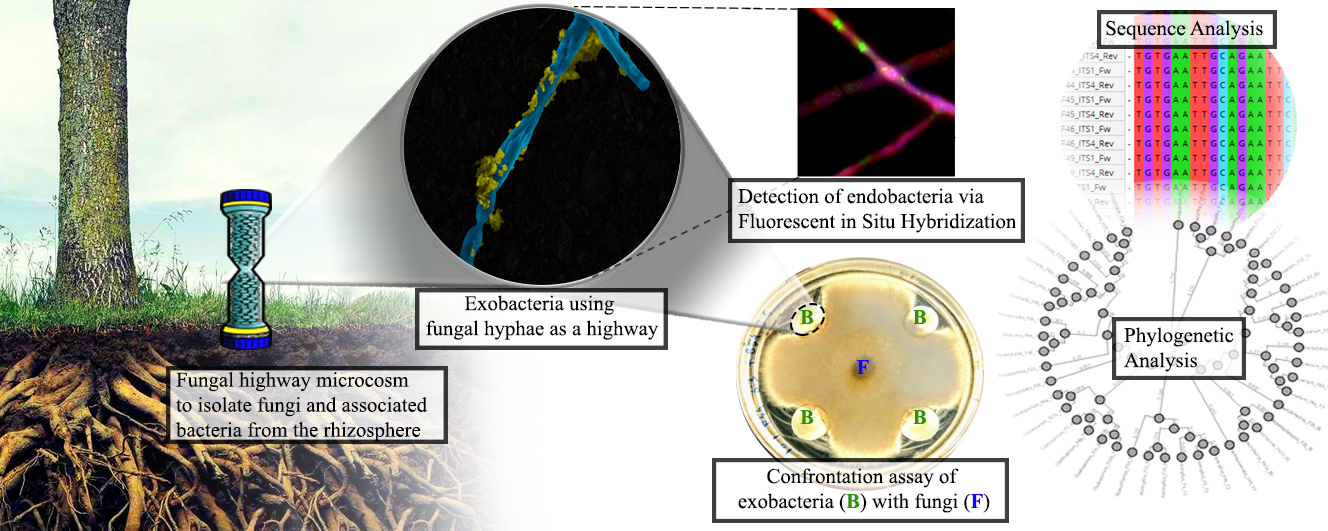
Bacteria have been described to colonize and use the fungal hyphae as highways to translocate in unsaturated soil matrices [51, 52]. Recently, a microcosm fungal highway column was developed as a unique isolation technique that allows bacteria to be transported using the fungus hyphae and to experimentally obtain insights on the bacterial-fungal interactions occurring in the soil [44, 54]. In the present study, this device was placed in the soil in proximity to the roots of six different plants using four different plant-based media as attractant for the isolation of cultivatable fungi and their associated bacteria from plant rhizospheres. The setup of the microcosm requires that the fungus grow upward in the unsaturated microcosm column to colonize the agar on top of the column. In this setup, the bacteria would also have to interact directly with the fungi to get transported to the top of the column. Therefore, we hypothesized that in these highway columns the bacteria should only have a positive or neutral interaction with the fungus to travel to the top of the column.
3.1. Fungal isolation from highway column samplings: relationship with types of plants and media
The fungal highway columns yielded a total of 46 fungal isolates from six different plant rhizospheres using four different plant-based media (Table 1). Most of the fungal isolates were identified as belonging to known plant pathogen species. The most represented fungal genera found in the collection were Fusarium and Cladosporium, both common soil-borne pathogens largely associated with plants and common inhabitants of the soil microbial community [55, 56]. Other fungal isolates, also associated with plant diseases, were Alternaria [57, 58], Diaporthe, and its asexual state Pomopsis [59, 60]. Moreover, we were able to isolate Aspergillus niger and Aspergillus fumigatus, which are often present in soils [61, 62], and Didymella, known for its pectolytic activities against plant cell walls [63]. We also isolated fungi often described as endophytes, such as Pestalotiopsis [64–69] and Plectospaerella [70], and non-pathogenic slow-growing mold, such as Stachybotrys [71] and Kalmusia. The latter is a fungus commonly associated with the soil crust [72]. In summary, all the fungal isolates have been previously described to be part of the soil microbial community.
Additional investigation involved the identification of possible relationships between plant type and the fungal isolates. For this purpose, we grouped the plants based on their tree or bush aspect and described them as tree sites and bush sites. In the case of the tree sites, they were divided into T1, T2, and T3, which corresponded to Citrus sinensis (Orange tree), Diospyros kaki (Persimmon tree), and Cycas revoluta (Cycad), respectively (Table 1). The Citrus sinensis (T1) was characterized by the fewest recovered fungal isolates (Fig. 2), i.e. one from Sorghum grain agar and one from Potato-carrot agar. Similar results and low fungal yield was previously observed in another study with the same type of tree [73]. These authors described that the principal taxa of fungi associated to Citrus sinensis was Basidiomycota, a generally slow growing phylum of fungi, and consequently difficult to isolate with conventional techniques of soil mycology [74]. This location was the only site where the genus Plectosphaerella was isolated (Additional file 1 - Supporting information). Plectosphaerella is a plant pathogen previously isolated from nine different plant genera [75–77] but never from Citrus sinensis. Kalmusia was another plant pathogen isolated that, as with Plectosphaerella, had never been described to be associated with orange trees. Among all the locations studied, the largest number of fungal species were obtained in the rhizosphere of Diospyros kaki (T2) (Fig. 2). This result may be explained by the natural decay of fallen persimmon fruits, which increased the nutrient content in the soil [78, 79]. Such a finding is also corroborated by the fact that the majority of the fungi obtained in this location were saprophytes [80–82].
In the case of the bush sites, they were divided into B1, B2, and B3, which corresponded to Ilex vomitoria (Yaupon), Myrica cerifera (Wax myrtle), and Buxus sempervirens (Boxwood), respectively (Table 1). Among all the bush locations, the number of fungi retrieved from these locations ranged from five, coming from the Buxus sempervirens (B3), to nine isolates from Ilex vomitoria (B1) (Fig. 2). Also, Ilex vomitoria (B1) was the only location with the presence of the fungus Diaporthales (Fig. 3). This fungus is a known plant pathogen commonly associated with various plants [59, 83]. The overall comparison of the bush and tree sites showed that the bush sites presented a higher diversity of fungal isolates (Fig. 3 – Supporting Information). In general, both sites seemed to share a core of four fungi commonly known as plant pathogens and soil inhabitants, namely, Pleosporales, Fusarium, Aspergillus and Cladosporium [55, 56, 61] (Fig. 3).
In addition to the types of plants, we determined the effect of different plant-based media on the diversity of fungi collected for each plant type. The results showed a noticeable difference in the culture collections, confirming previous findings on the effect of different media on the isolation of fungi [84–86]. Cornmeal agar recovered the highest number of fungal isolates, while oatmeal agar recovered the lowest (Fig. 2). Furthermore, we found at least one common taxon i.e. Fusarium and Pleosporales, in both tree and bush locations, as well as in all growth media, except for Potato-carrot. This finding suggests that these two taxa are ubiquitous in different plant rhizospheres.
Due to the intrinsic specificity of the nutrient requirement of the different microorganisms [87] and the fact that plant-based media do not have a defined chemical composition, it is difficult to draw a conclusion related to the effects of the media effects. However, we could conclude that none of the media used in this experiment can be defined as "optimal" considering the different number of fungi and bacteria obtained in the different media and different sites. Still, the community of culturable fungi was influenced by a combination of plant coverage types and the type of media used as a fungal attractant.
Table 1
Number of fungal and bacterial isolates obtained with the different media (Oatmeal Agar, Cornmeal Agar, Sorghum grain Agar, and Potato-carrot Agar) in six different types of rhizospheres. The area code in the table corresponds to the type of plant and was assigned as tree (T) and bush (B).
|
Area code
|
Description
|
Oatmeal
Agar
|
Cornmeal
Agar
|
Sorghum grain
Agar
|
Potato-carrot
Agar
|
|
Fungal
Isolates
|
Bacterial
Isolates
|
Fungal
Isolates
|
Bacterial
Isolates
|
Fungal
Isolates
|
Bacterial
Isolates
|
Fungal
Isolates
|
Bacterial
Isolates
|
|
T1
|
Citrus sinensis (Orange tree)
|
|
|
|
|
1
|
1
|
1
|
|
|
T2
|
Diospyros kaki (Persimmon tree)
|
2
|
|
6
|
5
|
5
|
5
|
3
|
12
|
|
T3
|
Cycas revoluta (Cycad)
|
2
|
|
|
|
4
|
1
|
|
2
|
|
B1
|
Ilex vomitoria (Yaupon)
|
3
|
|
6
|
|
|
|
|
4
|
|
B2
|
Myrica cerifera (Wax myrtle)
|
3
|
|
3
|
5
|
2
|
3
|
|
8
|
|
B3
|
Buxus sempervirens (Boxwood)
|
2
|
|
2
|
|
|
|
1
|
5
|
3.2. Linking fungal and bacterial isolates from the same columns
The isolated fungi and associated exo-bacteria carried along the columns with the fungal hyphae were further investigated. A total of 51 exo-bacterial isolates were successfully collected with serial transfers on plant-based media with fungicide (Fig. 4, Fig. 5). The most predominant phyla isolated were Firmicutes (57%), followed by Proteobacteria (29%) and Actinobacteria (16%). These phyla are the most prominent endophytic representatives described to be associated with diverse plants, such as Arabidopsis thaliana [89], Setaria italica (Foxtail millet) [90], Glycine max (soybean), [91] and Panicum virgatum (switchgrass) [92]. These results confirm the capacity of this isolation technique to obtain a similar rhizosphere microbiome to other techniques.
Among all locations, Bacillus was the most common isolate, followed by other well-known soil bacteria: Acinetobacter, Ensifer, Pseudomonas, Microbacterium, Paenibacillus, Rhizobium, and Stenotrophomonas. Most of these bacterial isolates are known to either act as plant growth promoters [93–104] or as biocontrol agents against plant pathogens [105–107].
Similar to the finding from the fungal isolates, Diospyros kaki (T2) presented the most exo-bacterial isolates, while Citrus sinensis (T1) presented the least. Among the tree rhizospheres, the total number of different bacterial isolates retrieved ranged from 22 associated with Diospyros kaki, to 1 with Citrus sinensis (Fig. 4). Interestingly, from the latter location, we were able to retrieve Pseudorhodoferax, a bacterium, that from the best of our knowledge, has never been described as being associated with the fungus Kalmusia.
In the bush sites, Myrica cerifera (B2) was characterized by the highest percentage of isolates ascribed to the genus Bacillus (62.5%). This genus was retrieved from almost all locations, except from Citrus sinensis (T1) and Cycas revoluta (T3) (Fig. 5).
Despite the small number of locations and isolates, a positive correlation was observed between the number of fungal and bacterial isolated in the different sites (Pearson's r2 = 0.84) (Fig. 7 – Supporting Information). Additionally, a positive correlation between the diversity indices of fungal and bacterial isolates were obtained from the different locations (Pearson's r2 = 0.76) (Fig. 8 – Supporting Information). From this data, we hypothesized that a species-specific partnership between fungus and bacterium could be occurring. However, the possibility of non-specific interactions cannot be excluded considering the co-occurrence of multiple bacteria and fungi retrieved in the columns. These results suggest that the abundance and diversity of the retrieved fungi should be able to directly affect the capacity to represent and recover the associated bacterial community. He et al. reported a similar result where a positive correlation was described when they studied the possible presence of a linkage between diversities of plants, fungi, and bacteria [108].
3.3. Bacterial-fungal interactions: reconstruction of the microbial associations in the soil
The positive linear correlation between fungal diversity and associated bacteria observed earlier suggested the need for a deeper understanding of the types of interactions between fungi and their associated bacteria. As mentioned earlier, we hypothesized that only positive or neutral interactions with fungi would have allowed the translocation of exo-bacteria via the fungal hyphae. Therefore, co-isolated bacteria and fungi in the same locations and media were further investigated via the confrontation assay (Fig. 6) in order to reconstruct the type of partnerships that were occurring in the soil.
As hypothesized, some of the bacteria isolated from the same location and growth medium displayed species-specific associations and neutral/positive interactions with the co-isolated fungus. This was the case for Agrococcus, Brevibacillus, and Exiguobacterium, which displayed positive or neutral interactions only in the presence of Didymella. A similar pattern was observed with the bacteria Cellulomonas associated with the fungus Staphylotrichum coccosporum and Stenotrophomonas with Aspergillus fumigatus. However, contrary to our initial hypothesis, in some cases different bacterial isolates belonging to the same species displayed different types of interactions with the same fungal isolate, suggesting that the bacterial-fungal interactions are not necessarily species-specific but rather strain-specific. (Additional file 4 - Supporting information). For instance, in the present study, we observed that some Bacillus isolates presented both neutral and antagonistic relationships with different fungal isolates of the same genus. One example was the association of Fusarium and Bacillus, where we recorded both negative and neutral interactions (Fig. 6). The negative interactions between Fusarium and Bacillus have also been observed and explained by others [109, 110].
The negative effects of bacteria toward fungi is typically a result of the bacterial production of antifungal secondary metabolites. For instance, as described by Mnif et al [111] lipopeptides produced by Bacillus display antifungal activity against F. solani. Another antifungal compound produced by different bacteria, including Pseudomonas, is 2,4-diacetylphloroglucinol (DAPG) [112]. DAPG is an active compound against various plant-pathogenic fungi, including Fusarium [113–116]. Besides the negative interactions observed between Fusarium and Bacillus, other studies also reported antagonistic interactions between Ensifer and the fungus Pleosporales, due to the production of alliinase, as a secondary metabolite for bacterial defense [113, 114, 117–119]. The present study also corroborates these previous findings that Bacillus, Ensifer and Pseudomonas present antagonistic patterns towards these fungal genera/species.
In addition to the negative interactions between Fusarium and Bacillus, this fungus also presented a neutral interaction with Rhodococcus. This interaction, however, was not species-specific since this bacterium also presented neutral interactions with other fungi. To further understand the nature of the neutral interactions observed in the present study, scanning electron microscopies were obtained. A representative image of the neutral interaction between Fusarium and the bacterium Rhodococcus is presented in Fig. 7. The results showed that the bacteria displayed the capacity to grow on top of the hyphae (Fig. 7). This type of finding was also reported by others [44]. However, it is important to point out that except for a few documented associations, such as the co-existence of A. fumigatus and Stenotrophomonas as a hyphae biofilm, [120] there is a lack of literature about those types of interactions in natural environments. This study shows that bacteria are commonly associated with fungi in plant rhizospheres. Furthermore, this isolation technique using highway columns promises to shed light on the specificity and types of relationships between bacteria and fungi in plant rhizospheres.
3.4. Endo-hyphal microbiome and bacterial-fungal specificity
Like other taxonomic groups of eukaryotic organisms, fungi have also been reported to have established associations with bacteria as endosymbionts [28, 29, 92, 121–124]. To determine if any of our fungal isolates harbored bacterial endosymbionts within their fungal hyphae, we performed both qualitative and quantitative investigations of our fungal isolates. Initial qualitative determination of bacterial presence inside the fungal hyphae was determined via 16S rRNA gene signal using fluorescence in situ hybridization (FISH) imaging (Fig. 8) [53]. The FISH results confirmed the presence of bacteria inside the hyphae of selected isolates. Figure 8 shows an example of bacterial signatures observed inside the Didymella mycelium. At low magnification, the presence of the signal (Cyan) can be observed ubiquitously across the fungal hyphae, while at higher magnifications bacteria can be observed within the hyphae. Innate fluorescence of the fungi was investigated to determine whether the fungi exhibited any fluorescent artifacts. Fusarium (F54) was also observed to exhibit internalized bacterial signals, which was to be expected since this fungus exhibited one of the highest bacterial loads determined by quantitative real time PCR (Additional file 1 - Supporting Information).
After confirming the presence of endosymbionts in the fungal isolates, 16S rRNA gene sequencing of these endosymbionts were done. The taxa of endobacteria present in each fungal isolated are listed in the Supporting information (Fig. 11, 12, 13, 14, 15, 16 – Supporting Information). On average, the fungal genera characterized by the largest numbers of endobacterial taxa were Pleosporales, followed by Fusarium (Fig. 13–14 – Supporting Information). This is the first study reporting the presence of endobacteria in Fusarium. Most of the endosymbiotic bacteria found in the genus Fusarium and in the order Pleosporales were composed of bacterial phyla commonly associated with the soil microbial community, such as Proteobacteria, Firmicutes, and Actinobacteria [125], suggesting that these bacteria probably have a mechanism to enter the fungal hyphae. Comparing exo-bacterial and endo-bacterial communities, we found that in most cases there was no intersection between these two microbial communities. Among the different exo-bacteria co-isolated with the fungi, only Bacillus, Microbacterium, Pseudomonas, and Stenotrophomonas were also found as endosymbionts. One of the most compelling examples is Bacillus spp. These species, despite having been associated as exo-bacteria with various fungal isolates (almost 70% of our collection), were present as endobacteria in only five fungi. More specifically, this genus was present in conjunction with the fungal taxa of Pleosporales, Didymella, and Neopestalotiopsis. This result validates the hypothesis that the endobacterial microbial community present in the fungi is not necessarily affected or linked to the exogenous microbial community. On the other hand, Pseudomonas and Stenotrophomonas were found to be present as both endo- and exo-bacteria, respectively, in 85% and 71% of their co-isolated fungi, suggesting that these genera could be horizontally transmitted from the fungus to the external environment and vice-versa [126].
The analysis conducted on the bacterial endosymbionts present in fungal isolates resulted in the observation that different genera of fungi present both a unique subset of bacteria and a shared core of five endosymbionts (Fig. 9) composed of isolates from Stenotrophomonas, Pseudomonas, Achromobacter, Candidatus Finniella, and Methylobacterium. This core of endosymbionts was previously documented as fungal endobacteria [121–123, 127, 128]. Among the core of endosymbionts, Stenotrophomonas [129, 130], Pseudomonas [131, 132] and, Achromobacter [133], they have also been described to be involved in nitrogen fixation in soil. Other endosymbionts, such as Bacillus [134], Bradyrhizobium [135], Novosphingobium [136], and Herbaspirillum [137], were also previously described to be participating in nitrogen fixation in soil [131–142]. Previous studies proposed that the bacteria could exploit the nitrogen resources of the fungus. The proposed mechanism involved the production of peptidases by the endophytic bacteria and consequent absorption of the resulting oligopeptides produced by their biodegradation [143, 144]. However, the identification of microorganisms, possibly capable of carrying out nitrogen fixation, present as fungal endosymbionts, indicate that there could be a more complex nutrient exchange pattern between the endobacteria and its host.
{kind=link}