We showed that T-cell responses to the FLS and N structural proteins of WT SARS-CoV-2 one-month following confirmed infection were similar in pregnant and postpartum black African women living with and without HIV who were naïve to Covid-19 vaccines. Virally suppressed WLWH on ART exhibited comparable polyfunctionality profiles compared to HIV-uninfected women, despite having a lower CD4+/CD8+ T-cell ratio. Even though all women were infected with the ancestral virus, Beta variant (B.1.351), or Delta variant (B.1.617.2) at the time of enrolment, cross-reactive T-cell responses were elicited to Omicron (B.1.1.529) and the ancestral virus to a similar degree, irrespective of HIV status.
The overall CD4+/CD8+ T-cell ratio was significantly lower in WLWH compared with HIV-uninfected women. The CD4+/CD8+ T-cell ratio is a biomarker that more accurately describes the overall risk of impaired immune responses compared to absolute CD4+ T-cell counts and HIV-1 viral loads [22]. A ratio above 1.0 is considered normal, but can be influenced by various factors such as age, sex, and ethnicity [22, 23]. A lower CD4+/CD8+ T-cell ratio is generally predictive of adverse clinical outcomes [23], whereas ratios above 2.4 have been associated with increased risk of fatal outcomes during acute Covid-19 [24]. However, the CD4+/CD8+ T-cell ratio appears to remain constant during SARS-CoV-2 infection in PWH compared with HIV-uninfected individuals in whom the ratio is increased [25].
We did not observe a difference in the CD4+/CD8+ T-cell ratio between pregnant and postpartum women, with only pregnant WLWH exhibiting a lower ratio compared with HIV-uninfected pregnant women. In addition, all pregnant, postpartum, and non-pregnant women living with HIV had ratios below 1.0. The lower ratios can be ascribed to a greater proportion of CD3+ T cells with a CD4+ phenotype or more likely, a greater proportion of CD3+ T cells with a CD8+ phenotype which did not normalize despite ART [26]. While the duration of ART can be considered a confounder, it is important to mention that information on ART duration was not reported for the participants enrolled in our study.
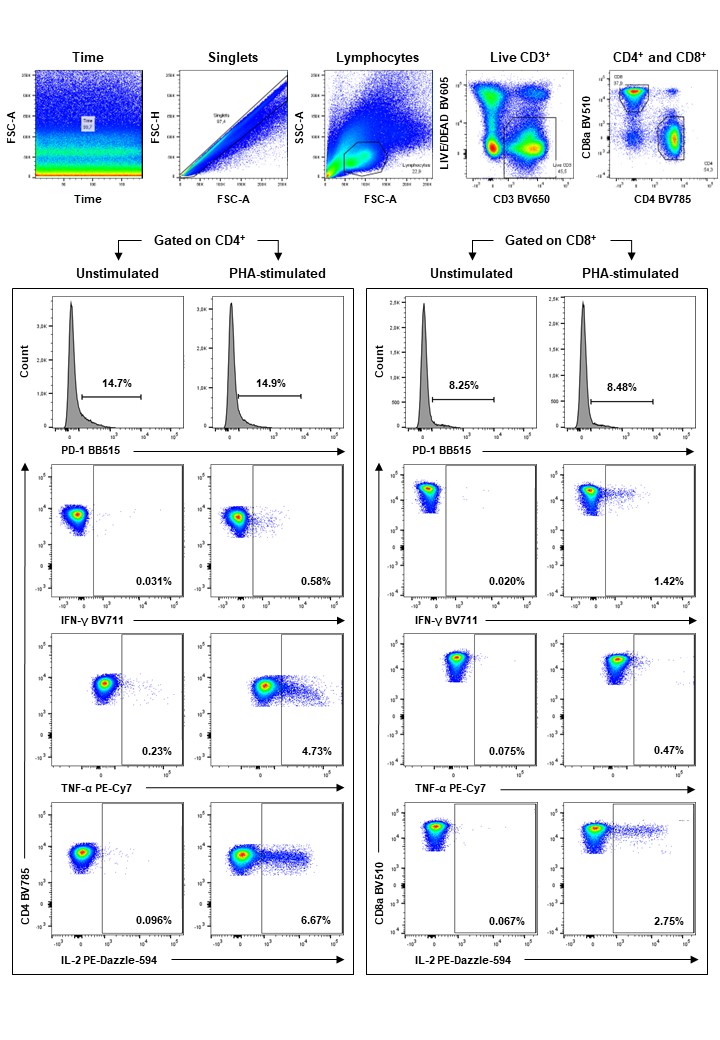
We demonstrated that higher frequencies of CD4+ T cells express PD-1 (CD279) compared with CD8+ T cells in women living with and without HIV one month post positive SARS-CoV-2 diagnosis. PD-1 is a down-regulator of immune responses and suppressor of T-cell inflammatory activity [27]. During SARS-CoV-2 infection, PD-1 is expressed on both CD4+ and CD8+ T-cell subsets, however, is significantly increased on CD4+ T cells during acute Covid-19 [28, 29]. Our results support the expression of PD-1 on both CD4+ and CD8+ T cells one month post infection among both HIV-uninfected women and WLWH, whose SARS-CoV-2 viral loads were similar at the time of diagnosis. The increased proportions of PD-1-expressing CD8+ T cells in WLWH relative to HIV-uninfected women is likely a consequence of long-term chronic HIV-1 infection rather than of acute SARS-CoV-2 infection alone [30]. As all participants were recruited as incidental SARS-CoV-2 infections, we could not assess differences between the study groups in the anticipated increase in PD-1-expressing T cells because of acute SARS-CoV-2 infection. Immune activation persists in PLWH despite ART, contributing to a more exhausted immune profile as evidenced by increased proportions of PD-1 expressing CD8+ T cells [30–32].
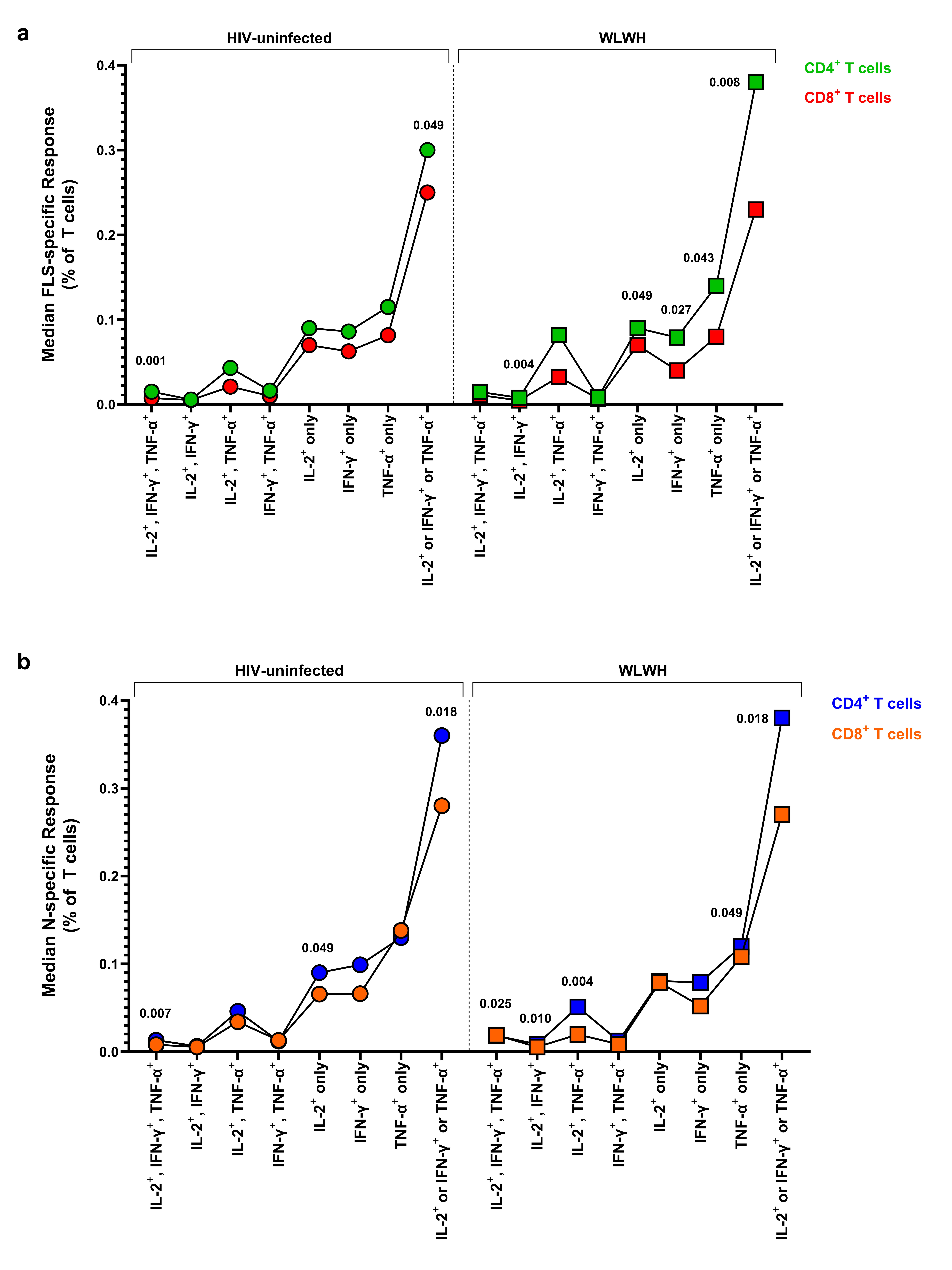
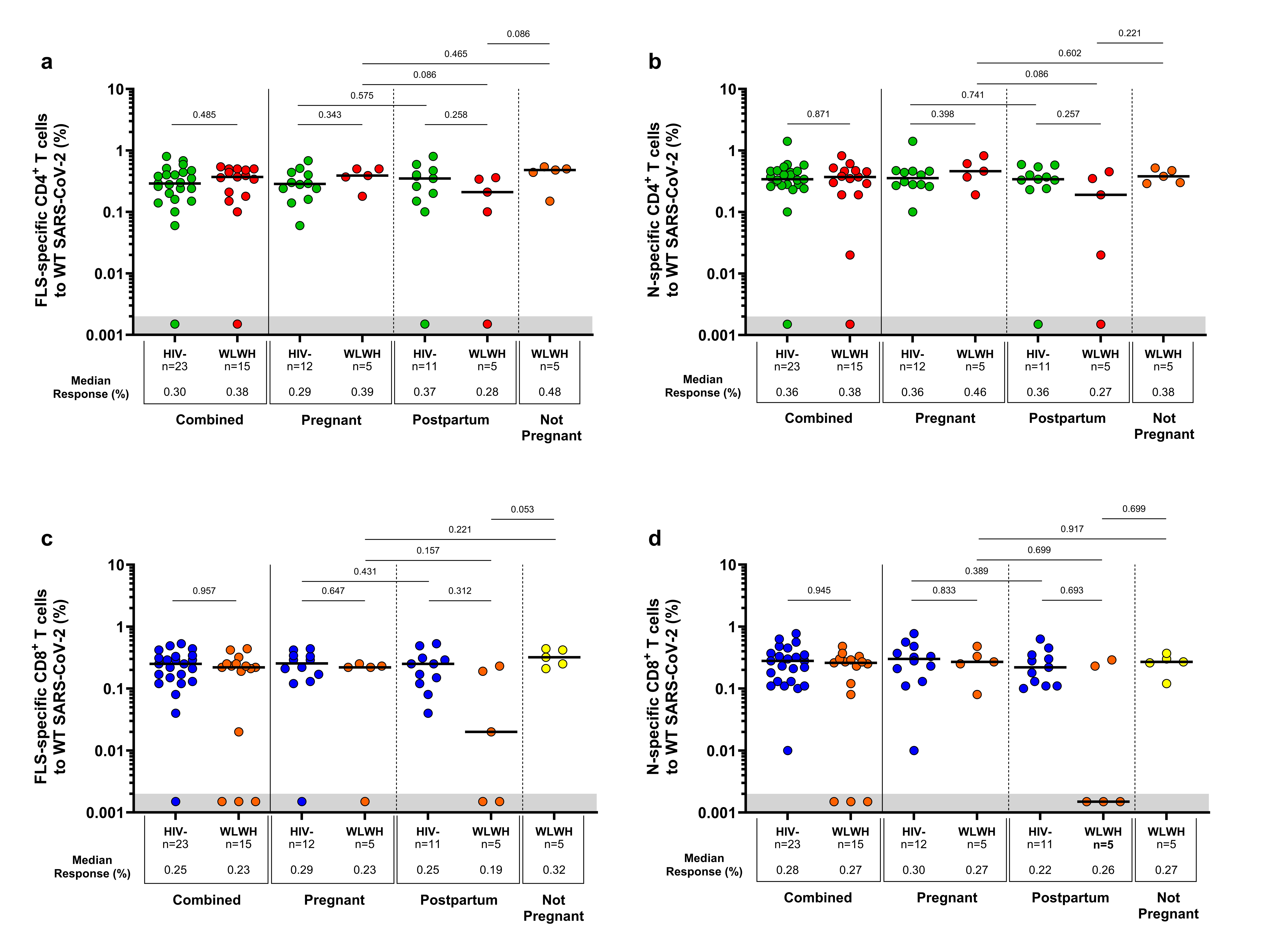
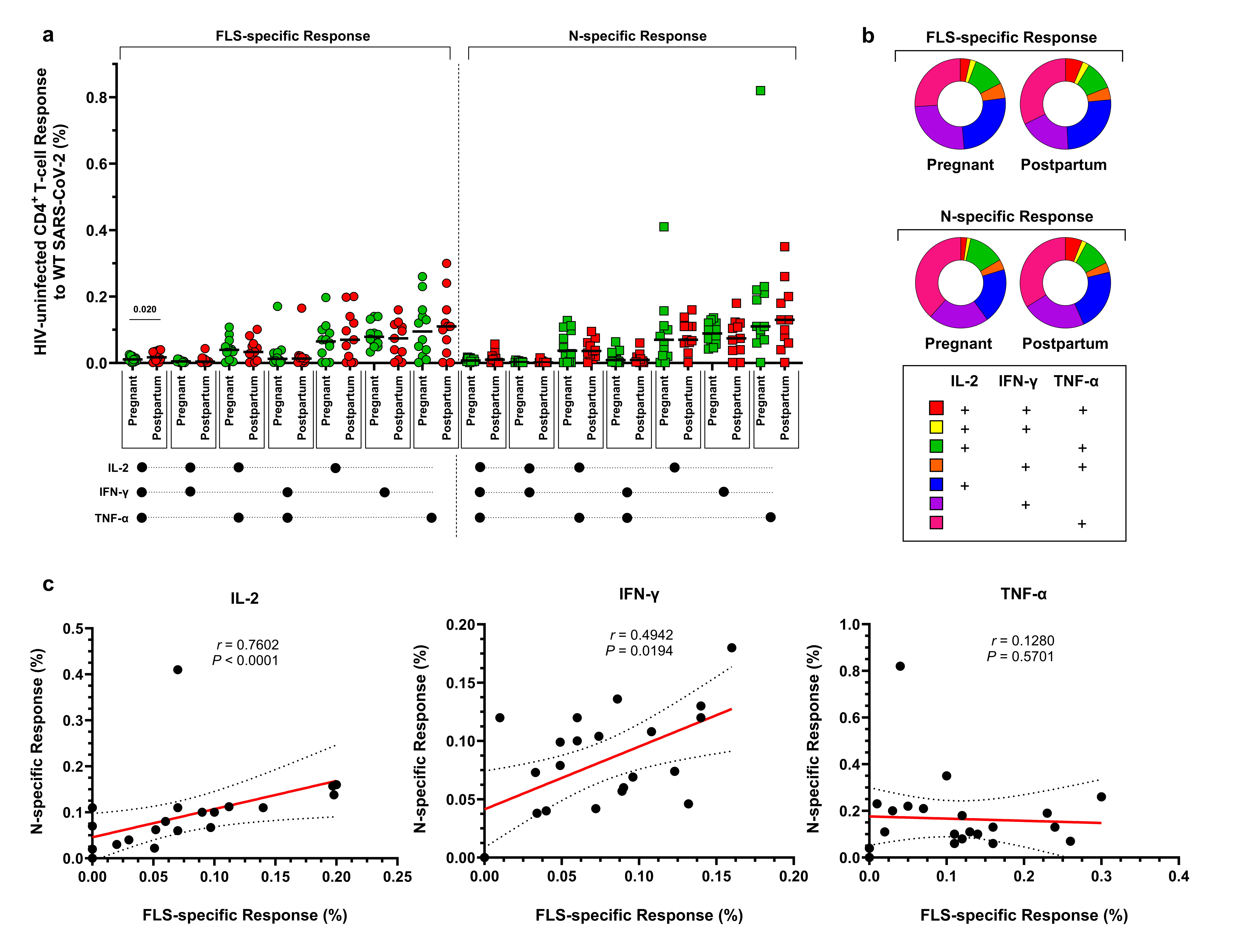
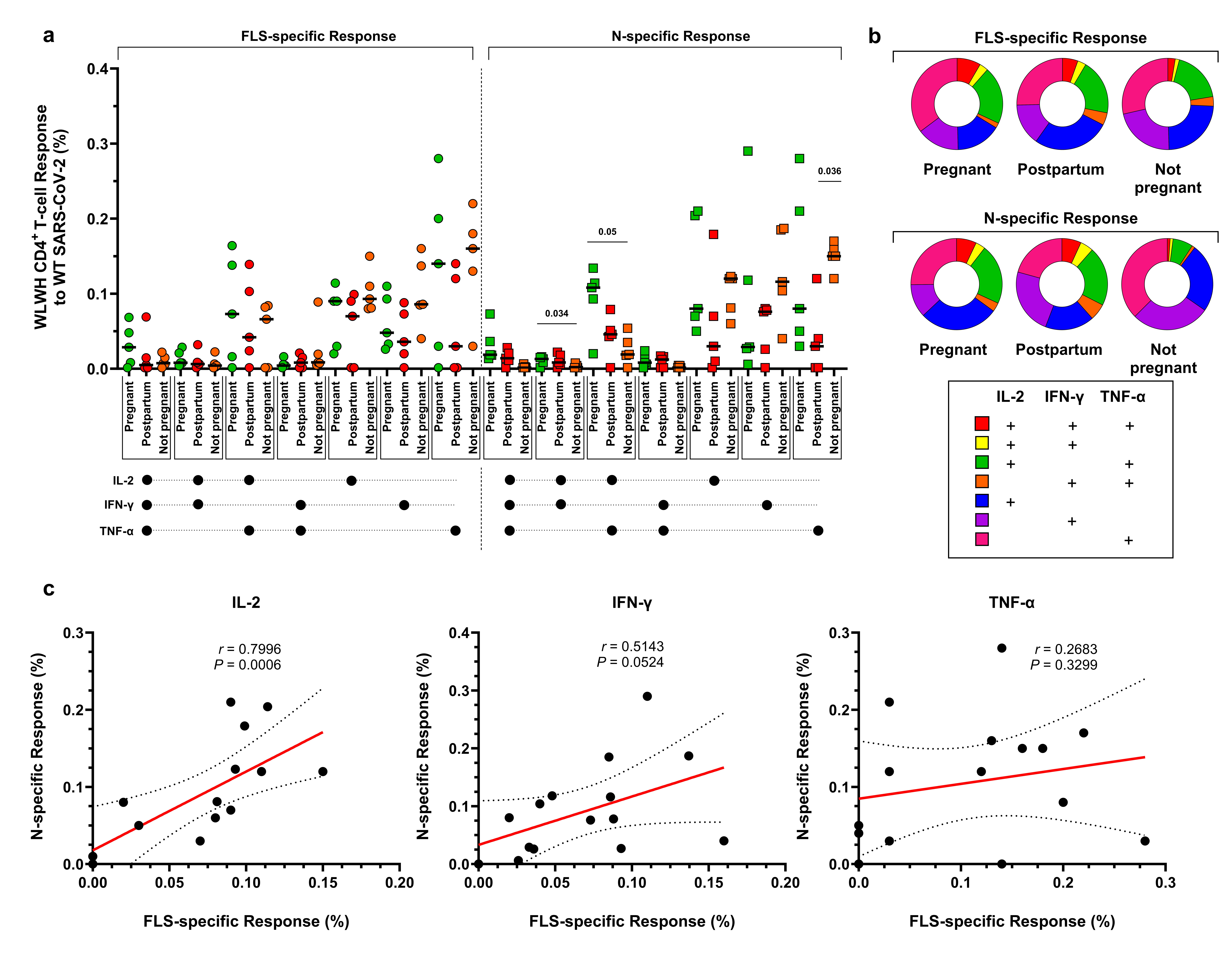
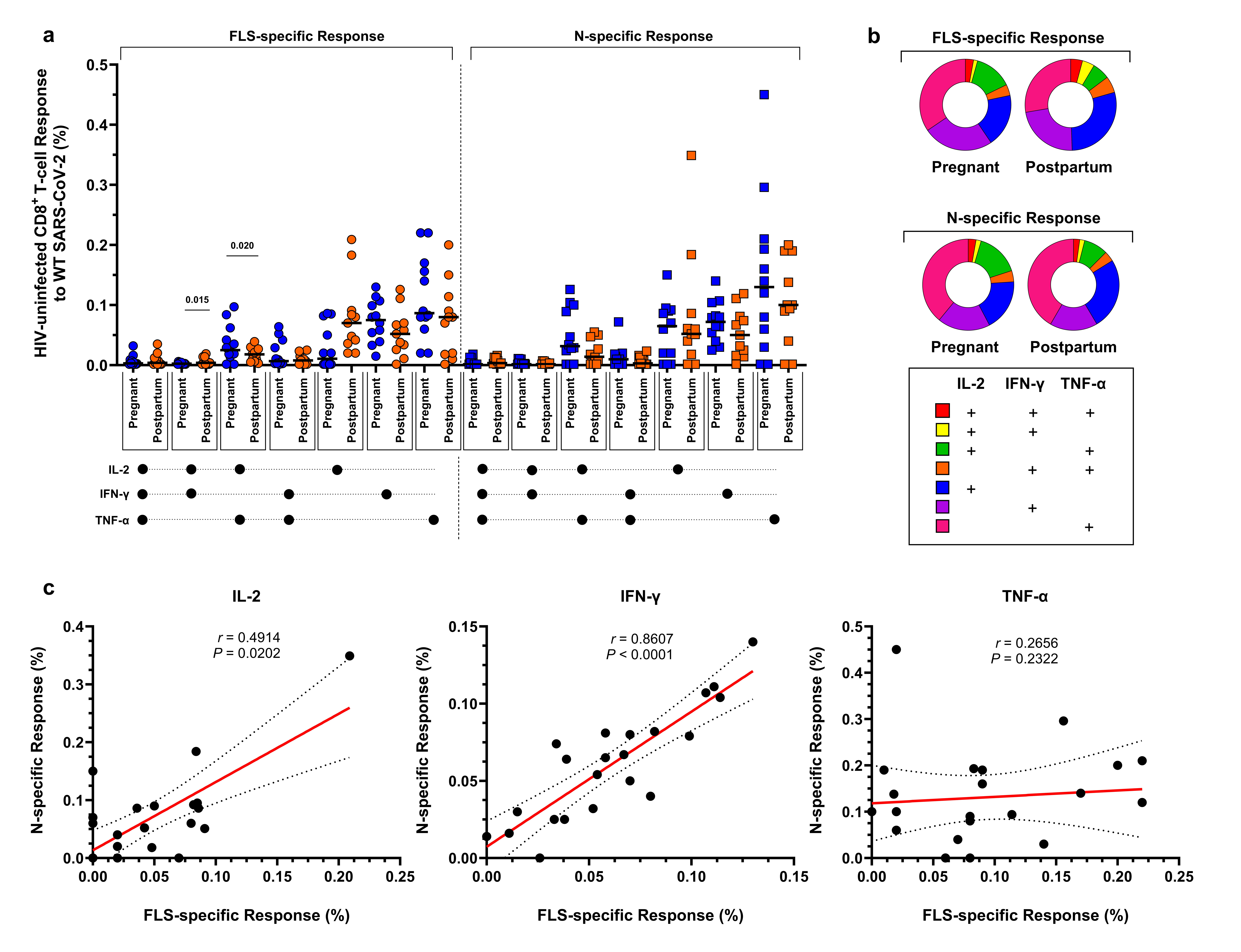
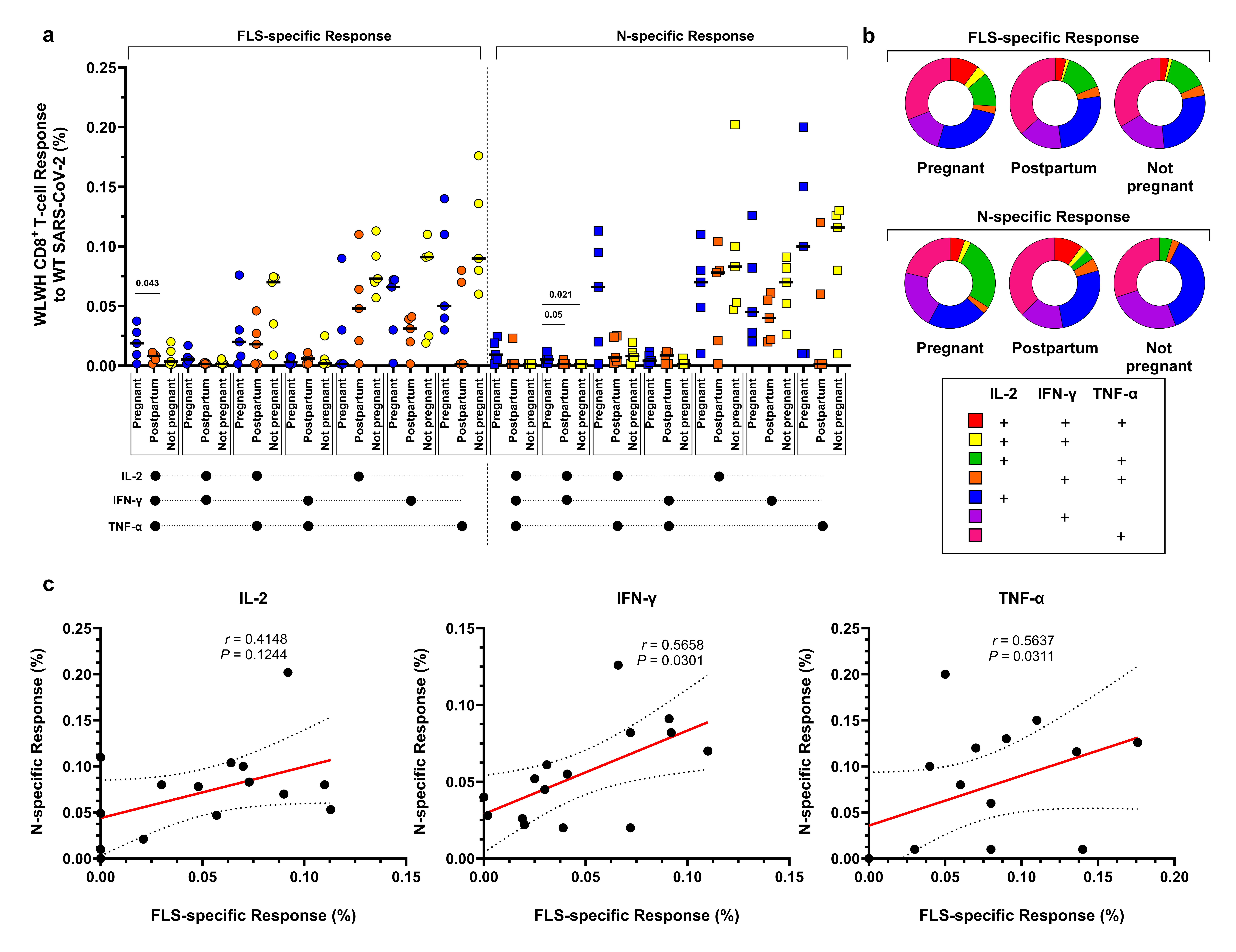
T-cell responses directed at the FLS and N proteins of WT SARS-CoV-2 one-month post infection showed a modest correlation with each other, and were also similar between pregnant and postpartum women living with and without HIV. Our findings of similar T-cell responses irrespective of HIV infection status, corroborate the observations in a previous study from our group in similarly healthy African adults on ART who received the ChAdOX1 nCoV-19 (AZD1222) vaccine [33]. Other studies have similarly indicated that the SARS-CoV-2-specific CD4+ T-cell response magnitude outnumbers the CD8+ T-cell response magnitude [29, 34]. However, 15-mer peptide antigens may have underrepresented the contribution of SARS-CoV-2-specific CD8+ T-cell responses, since they are optimal for HLA class II binding and have been reported to capture on average only 77% of CD8+ T-cell responses compared to shorter peptides [35].
Reduced numbers of CD4+ T cells in PWH have been linked to suboptimal SARS-CoV-2-specific humoral and cell-mediated immune responses [28]. Not only do SARS-CoV-2-specific CD4+ T cells play a critical role in the control and resolution of acute Covid-19 [36–38], but they also assist CD8+ T cells and B cells with long-term immunity [39, 40]. SARS-CoV-2-specific T-cell responses in black Africans living with uncontrolled HIV infection, have previously been linked to impaired T-cell immunity with a lower T-cell response magnitude, reduced polyfunctionality, as well as diminished cross-recognition of CD4+ and CD8+ T-cell subsets [17, 41]. We show that WLWH on ART exhibited comparable polyfunctionality profiles compared with HIV-uninfected women, despite having a lower CD4+/CD8+ T-cell ratio and increased proportions of the CD8+ T-cell subset.
We could not identify any discernible patterns in the polyfunctionality response profiles among pregnant, postpartum, and non-pregnant WLWH, but we observed comparable T-cell response profiles in HIV-uninfected women. During pregnancy, there is an upregulation of innate immune responses, as well as a natural shift from Th1 immune responses to Th2 responses [42]. The postpartum period may reflect aspects of pregnancy-associated alterations for an unknown period and the return to a pre-pregnant immune state is not well characterized, although it may take up to 12 months [43]. This may explain why we observed similar T-cell responses between pregnant and postpartum women living with and without HIV.
Lastly, we demonstrated robust CD4+ and CD8+ T-cell responses that cross-react with Omicron to a similar degree compared with WT SARS-CoV-2, irrespective of HIV-status. Our results are consistent with other studies investigating potential T-cell escape by Beta, Delta, and Omicron variants [29, 44, 45]. Despite spike-specific mutations in SARS-CoV-2 variants, which may diminish the cross reactivity of certain T-cell epitopes, broad T-cell responses developed following SARS-CoV-2 infection in response to immunodominant epitopes located across the viral proteome [46, 47]. Therefore, T-cell immunity is mostly conserved and peaks between days 14–30 post infection [17, 29, 45].
Our study had some limitations. We only characterized SARS-CoV-2-specific T-cell immunity by focussing on the ability of T-lymphocytes to produce IL-2, IFN-γ, and TNF-α. These cytokines are characteristic of a Th1 immune response. Since we included pregnant and postpartum women living with and without HIV, the investigation of Th2 immune responses would have provided additional information about the immunological landscape [48, 49]. Furthermore, we had limited numbers of WLWH at one-month post SARS-CoV-2 infection, which limits the power of our analyses and the interpretation of the results. This may explain why we were unable to identify any apparent patterns in polyfunctionality profiles between pregnant, postpartum, and non-pregnant women living with HIV. We did not perform viral genome sequencing following confirmed SARS-CoV-2 infection to confirm the identity of the variant. The identity of the SARS-CoV-2 variants were inferred solely by temporal association in relation to the enrolment periods that coincided with variant-specific Covid-19 waves in South Africa. The possibility remains that seasonal human coronaviruses or other SARS-CoV-2 variants could have been responsible for the detected T-cell responses, not only WT SARS-CoV-2, Beta variant, or Delta variant. Lastly, since these women were tested for SARS-CoV-2 infection during routine antenatal care, positive cases were purely incidental detections. We do not know exactly when the infections occurred and how the results might have differed for women with severe Covid-19.
In conclusion, SARS-CoV-2 infection induced comparable T-cell responses and polyfunctionality profiles between pregnant and postpartum women living with and without HIV who were naïve to Covid-19 vaccines. Since T-cell immunity develops in response to immunodominant epitopes located across the viral proteome, our investigations of only two immunodominant structural proteins are reassuring to HIV-burdened low- and middle-income settings where a significant proportion of the population were infected before Covid-19 vaccination. Additionally, cross-reactive T cells are elicited against Omicron and WT SARS-CoV-2 to a similar degree in women infected with the ancestral virus, Beta variant, or Delta variant; which further confirms the overall preservation of T-cell immunity despite variant-specific spike mutations.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}