The genus Angiostrongylus Kamensky, 1905, contains the zoonotic nematode parasites of vertebrates commonly known as “lungworms” [1]. Several species in this genus have been reported to be human pathogens, including A. cantonensis and A. costaricensis, while A. mackerrasae and A. malaysiensis are considered potential human pathogenic parasites [2–5]. Among these species, A. cantonensis is a well-known pathogen that causes angiostrongyliasis eosinophilic meningitis in humans [6], over 2,800 cases of which have been reported worldwide, with a particularly high incidence in Thailand [7]. Humans are accidental hosts and are infected by eating undercooked snails, slugs, paratenic hosts, and contaminated fresh vegetables containing the infective third-stage larvae [8]. The clinical symptoms can range from headaches to coma depending on the number of parasitic larvae [2, 8–10]. Many species of terrestrial and freshwater snails, for example, Achatina fulica, Cryptozona spp., Pomacea spp., and Pila spp., are reported to be critical vectors of A. cantonensis [11–13]. Global parasite transmission frequency has been promoted by the widespread distribution of some invasive vector species, for example, A. fulica and Pomacea spp. [13–15]. The ability of the parasite to spread throughout the world may also be influenced by its natural definitive host, a rodent that inhabits areas of human urbanization [1, 6, 16].
Among species of the genus Angiostrongylus, a cryptic species of A. cantonensis was recently correctly described as A. malaysiensis [17, 18]. Previously, A. malaysiensis was known as the Malaysian strain of A. cantonensis [1, 5]; however, its pathology differed from the typical pathology of A. cantonensis through its ability to migrate to the central nervous system of the monkey host but not cause death [1, 19]. The strain was known to immunize monkeys against the lethal effects of a high number of A. cantonensis larvae. Subsequently, the Malaysian strain was species delimited as A. malaysiensis [5, 20].
The adult stages of A. cantonensis and A. malaysiensis can be morphologically discriminated by the structure of the bursal rays of males and the minute protrusions at the posterior of females [21]. The third-stage larvae of A. cantonensis are described as using the mellifluous “Q”-movement, which clearly distinguishes them from free-living nematodes [22]. However, differences between the morphological characteristics of the larval stage A. cantonensis and A. malaysiensis have not yet been described.
Previously, the morphological variations between these two species were never considered, and their similarities were the cause of the misidentification. A. cantonensis was known as the predominant species in the genus of Angiostrongylus, with distribution throughout many countries in Indochina [23]. In 2016, Rodpai et al. revealed that A. cantonensis is widely distributed in Thailand, Lao PDR, Cambodia, and Myanmar, and its distribution broadly overlapped that of A. malaysiensis [23]. Dusitsittipon et al. discussed the incongruence between the morphological and molecular identifications and found that most of the misidentified specimens were from Thailand [17, 24].
An under-estimation of the prevalence of A. malaysiensis has occurred because of the difficulty in the morphological identification of adult and third-stage larva. Several polymerase chain reaction (PCR)-based identification techniques were developed, but all were aimed at increasing the sensitivity and specificity of A. cantonensis detection in blood, the peripheral tissues of wild Hawaiian rats, and cerebrospinal fluid (CSF) of patients [25–27]. No powerful techniques with high sensitivity and specificity have been developed for detecting and discriminating between A. cantonensis and A. malaysiensis.
A. cantonensis and A. malaysiensis can be distinguished using the mitochondrial cytochrome b (Cytb) gene as a genetic marker because of the robust phylogenetic relationship between the closely related species [17, 24]. This gene sequence shows sufficiently high genetic variation between species, and species-specific primers can be used to detect and discriminate between the two species with high sensitivity and specificity. The SYBR-green quantitative real-time PCR method (qPCR) was considered suitable for quantitative detection and species discrimination because it is cheaper than TaqMan qPCR, which requires a fluorescent probe. SYBR green qPCR has recently been used in many studies to detect, discriminate, and quantify species as, for example, in the discrimination and quantification of Leishmania in human samples and the discrimination of species and subspecies of Salmonella. The technique was also employed for the detection of Opisthorchis viverrini and Haplorchis taichui from human stool samples [28–30].
In this study, therefore, we designed species-specific primers to Cytb partial gene sequences, which have sufficiently high genetic variation between species to prevent cross-amplification [17, 24]. We aimed to develop a highly sensitive and specific method capable of quantitively detecting the third-stage larvae of A. cantonensis and A. malaysiensis in their intermediate host, Achatina fulica, and to determine the efficacy of the method for the detection of Angiostrongylus genomic material within the CSF of infected patients.
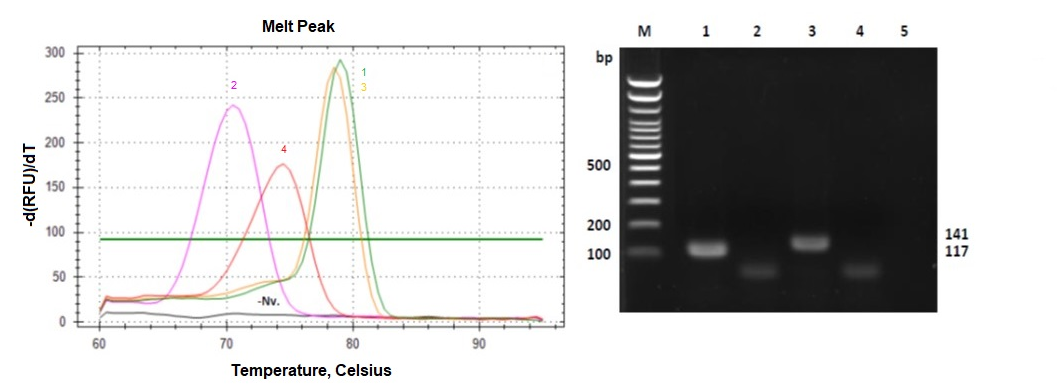{kind=link}
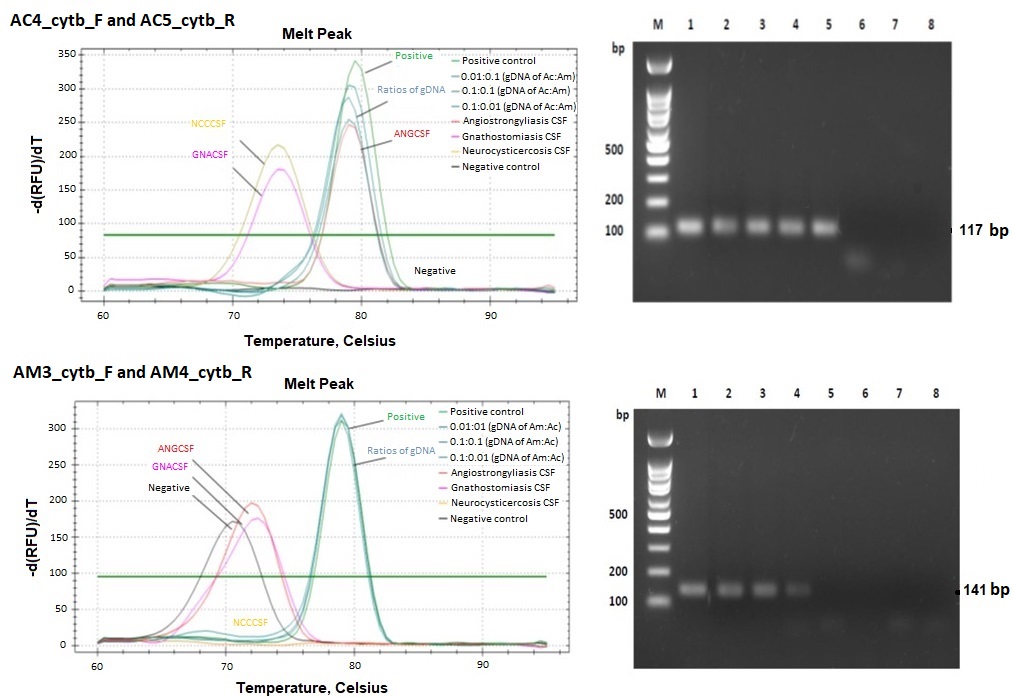{kind=link}