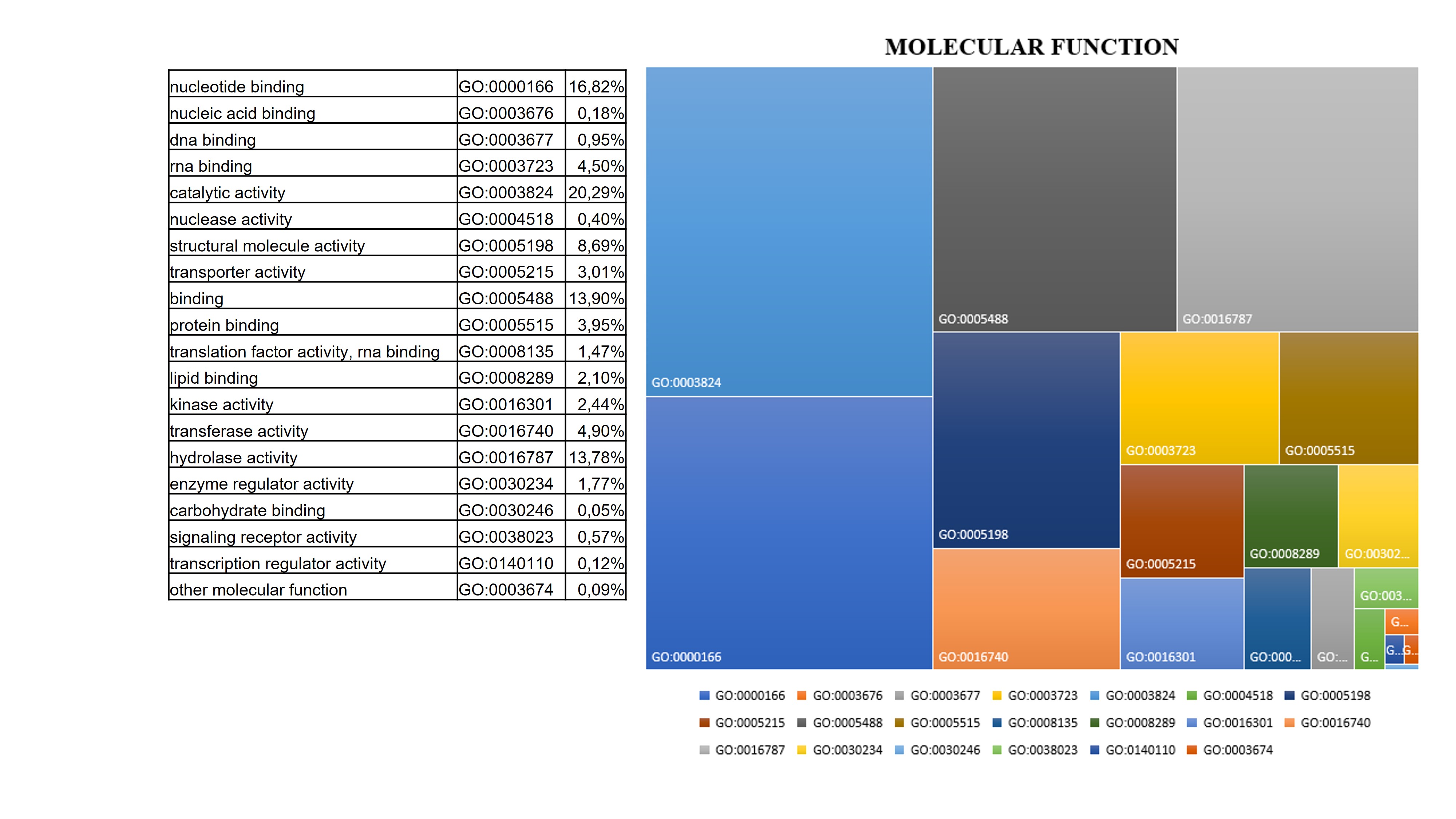
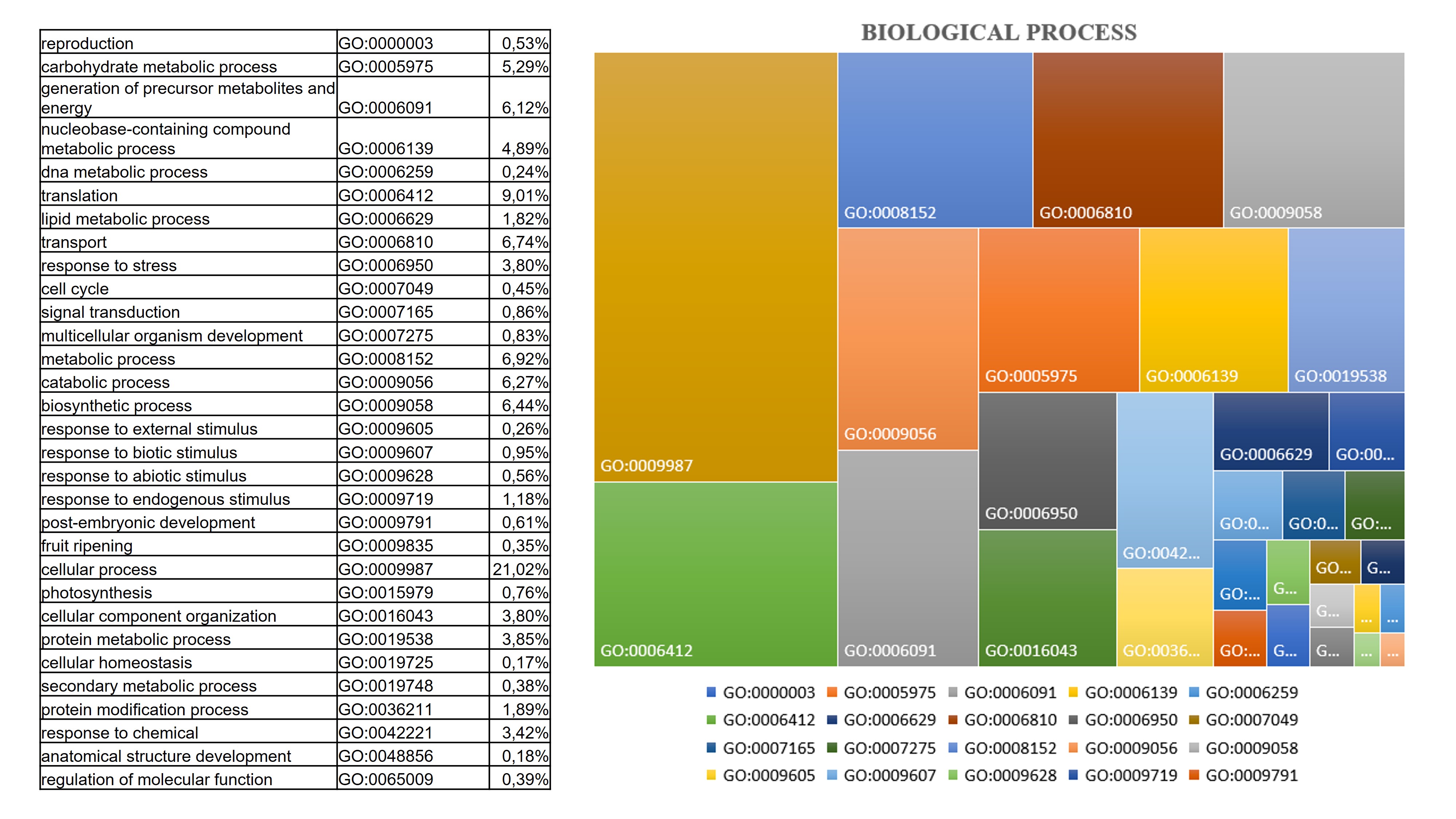
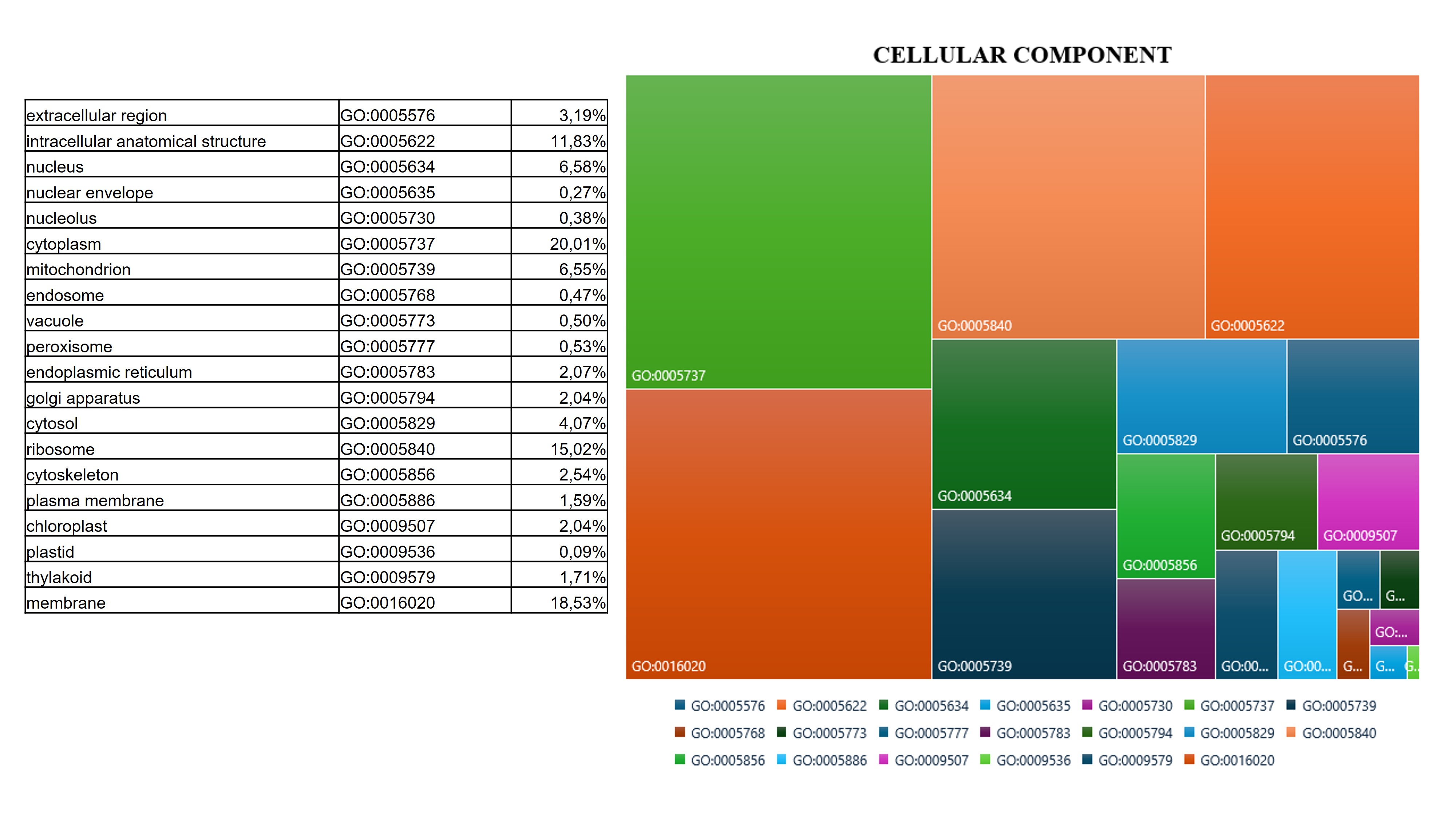
Although much information is available for other fruits belonging to the Rosaceae family regarding physiological and biochemical changes due to the ripening process, little is known about the postharvest ripening process of Cormus domestica (L) fruit. This small, ancient fruit, widespread in the Mediterranean, and closely connected to traditional and rural culture, is unfortunately at serious risk of cultural erosion due to the wider spread of intensive cultivation of fruits with wider economic value such as the apple or pear[19]. The ripening process is a genetically programmed, irreversible and highly regulated process involving changes in the fruit structure and chemical composition. Generally, the combination of physiological changes makes the fruit more palatable for consumption. The main alterations of a ripening fruit involve its colour change mainly due to the chloroplasts turning into chromoplasts and the accumulation of neo-synthesised pigments, softening of the pulp due to the alteration of the cell walls, sweetening due to the accumulation of sugars and the reduction of sour components, and the production of various metabolites that contribute to the attainment of aroma and flavour [13, 20–22]. Traditionally, C. domestica fruits are harvested fully developed, and eaten after a maturation period in a cool, dry and airy room in the dark for about a month until they reach a soft consistency. Only then will the fruit have lost its acrid, woody flavour and become very sweet and juicy. At this stage it is consumed as it is or used for the production of jams, jellies or spirits[17]. The pronounced colour change, usually observed during the fruit ripening [23] is not observed in C. domestica fruit, which, as shown by analyses of photosynthetic pigment content and in the sample photos shown, does not accumulate carotenoids during ripening, but rather takes on a brownish colour mainly due to the natural oxidative processes involving the fruit in post-harvest ripening[24], and as shown by the accumulation of enzymes involved in oxidative stress found in our samples. In addition to colour change, ripening includes an essential softening process due to hydrolytic processes resulting in the breakdown of cell wall polymers (cellulose, hemicelluloses and pectins) by wall enzymes that are essential for softening the fruit and improving its chewability[14]. One of the most abundant functional classes revealed by our proteomic analysis is that involved in cell wall changes. In fact, the results highlighted that the abundance of enzymes such as b-galactosidase and pectinesterase act on the pectic component of the cell wall, which is mainly responsible for the hardness of unripe fruits. Specifically, the role of β-galactosidase (β-Gal) in the apple ripening process has been studied and its involvement in modulating the firmness and the pectin content has been confirmed and appears to be induced by ethylene [25]. These enzymes, like others that act on hemicelluloses (XET) and non-enzymatic proteins such as expansin, accumulate abundantly in the fruit of C. domestica already in the early stages of post-harvest ripening. The enzymatic activity of the cell wall in the fruit of C. domestica makes it particularly soft in consistency, but this, together with the accumulation of sugars and oxidative processes, significantly reduces its shelf life [14]. Reactive oxygen species (ROS) are inevitably produced by metabolic pathways activated during the fruit ripening process. The ROS accumulation causes oxidative damage by reacting with nucleic acids, proteins and lipids. Simultaneously with their production and accumulation in plant tissue, plants activate the recovery system and produce enzymatic and non-enzymatic antioxidants such as superoxide dismutase (SOD), glutathione reductase (GR), peroxidase (POX), catalase (CAT), ascorbate peroxidase (APX), monodehydroascorbate reductase (MDHAR), dehydroascorbate reductase (DHAR), and antioxidant compounds such as ascorbate, glutathione, flavonoids, and tocopherols, which were found to accumulate in the C. domestica fruits under analysis and as in other fruits belonging to the Rosaceae family [11, 26]. The accumulation of antioxidant enzymes and soluble antioxidant compounds such as flavonoids has also been observed in the fruit of the service tree. In particular, the accumulation of quercetin-3β-D-glucoside, taxifolin, fustin and coumarin increase the nutraceutical power of this ancient fruit [27, 28], which has a general anti-inflammatory effect and can modulate the lipid peroxidation involved in various diseases such as atherogenesis, thrombosis, and carcinogenesis, especially at the habitual stage of fruit consumption to decay then 4 weeks after harvest. Ripening is a physiological process, highly regulated with pathways that are still not clearly defined and understood, and which interconnects at several levels with abiotic stress response pathways[20]. The post-harvest ripening process induced in C. domestica fruit the accumulation of Abscisic stress ripening protein (ASR), a protein induced by ABA, stress and ripening. Several studies have shown that ASR gene expression varies between species, organs and conditions, and appears to be involved in fruit ripening, although its mechanism of action is not yet well understood [29, 30]. Similarly, Remorin, a protein involved in the regulation of several physiological processes, shows a peak in abundance two weeks after service tree fruit harvest, and its contribution in regulating the ripening process is known from the literature. Other proteins involved in stress and energy metabolism such as Small Heat Shock Proteins (sHSPs) were found to be abundant in the ripening C. domestica fruit. These chaperones have been found in several proteomics studies related to abiotic and developmental stress response that have been characterised in several ripening fruits [31, 32]. During ripening, a complex regulatory mechanism shifts metabolism from starch synthesis to starch degradation and the accumulation of soluble sugars, mainly sucrose. This conversion is responsible not only for fruit sweetness, but also for providing energy for the metabolic processes taking place during ripening [33–35]. As expected, also in the post-harvest ripening process of the service tree fruit followed in this experiment, a marked conversion of starch to soluble sugars was observed and the levels of the enzyme sucrose synthase remain high until the end of the ripening process. The main soluble sugars in sorbola-like fruits such as apples or pears are sucrose, fructose and glucose, which is also particularly abundant in the sorbola stage of consumption; as are some sugar alcohols such as sorbitol [9]. Furthermore, metabolic analysis confirmed the gradual but significant accumulation of maltose and glucose as ripening progressed. In addition to sweetness, another important parameter that determines the organoleptic characteristics of the fruit is acidity, mainly due to the presence of organic acids accumulated in the vacuole. In apples and pears, the main ones are malic acid and citric acid, and the lowering of the degree of acidity is mainly due to the investment of these organic acids in the respiration process and the production of secondary metabolites. Citrate synthase and Malic enzyme accumulate in the service tree fruit during ripening, causing a reduction in malate and contributing to the decline in fruit acidity [36]. Several peculiar metabolites characterize the habitual stage of fruit consumption. Some of these, particularly abundant such as piptamine and resorcinol, are known for their antimicrobial and antifungal properties. In addition to firmness and sweetness, another important parameter characterising a ripe fruit is its olfactory characteristics. These depend on hundreds of volatile compounds that are produced differentially during the various stages of ripening and are responsible for aroma production. Volatile compounds vary widely in different fruits and are generally aldehydes, alcohols, ketones, lactones, esters and terpenoids. Because aroma is closely associated with phytochemicals that make fruit healthy, the health benefits for the consumer are not limited to the satisfying experience for the senses (taste and smell). Metabolomic analysis has allowed us to identify several compounds particularly abundant in the preferential eating stage of the C. domestica fruit that together provide the fruit with a peculiar aroma. Citral and limonene, in addition to inducing the jasmonic acid pathway to endow fruit with resistance to biotic stresses, confer fresh and citrus notes. The abundance of camphor, pulegone and borneol, on the other hand, adds intense minty notes, while benzophenone adds soft, fresh notes with touches of fruit and flowers. These compounds not only impart the final aroma to the fruit but also have antimicrobial and anti-inflammatory properties[13, 37, 38].
{kind=link}
{kind=link}
{kind=link}
{kind=link}