In the current study, we explored the use of the Nanostring nCounter technology and the predefined Human v3b miRNA Assay panel to identify dysregulated miRNAs in the pig brain, in response to a single psychedelic dose of psilocybin. To our knowledge this has not previously been done. We report miRNA-alterations related to acute (PSI-1-day) and prolonged (PSI-7-day) effects of psilocybin in the PFC and HIP of pigs. In total, across the PFC and HIP, we identified 14 and 4 dysregulated miRNAs out of 827 miRNAs in after 1 day and 1 week, respectively.
Generally, we observed greater miRNA-response to psilocybin in the PFC than in the HIP. A possible explanation might be the 5-HT2AR distribution in the brain, as the 5-HT2AR display high densities in the cerebral cortex whereas the densities in the HIP are very low28.
The miRNA-regulation observed after 1 day in the PFC generally reverted to baseline levels after one week. On the contrary, we observed the largest regulation in the HIP after 1 week. It can be speculated that this may be caused by psilocybin-induced modifications in the PFC, which stimulate 5-HT2AR-independent mechanisms in the HIP in a delayed manner.
Nine of the 18 miRNAs identified in our study have previously been associated with depression
miR-92a-3p, miR-98-5p, miR-99b-3p, miR-107, miR-125a-5p, miR-128-3p, miR-212-3p, miR-221-3p, and miR-485-3p have been identified as dysregulated in brain tissue or blood from rodent models of depression and depressed patients29, 30 (summarized in Table S4).
Interestingly, miR-107 and miR-212-3p, which were found differentially regulated in PFC using all 4 normalization methods, were two of the nine miRNAs.
miR-212-3p is upregulated 1 day after psilocybin administration in PFC
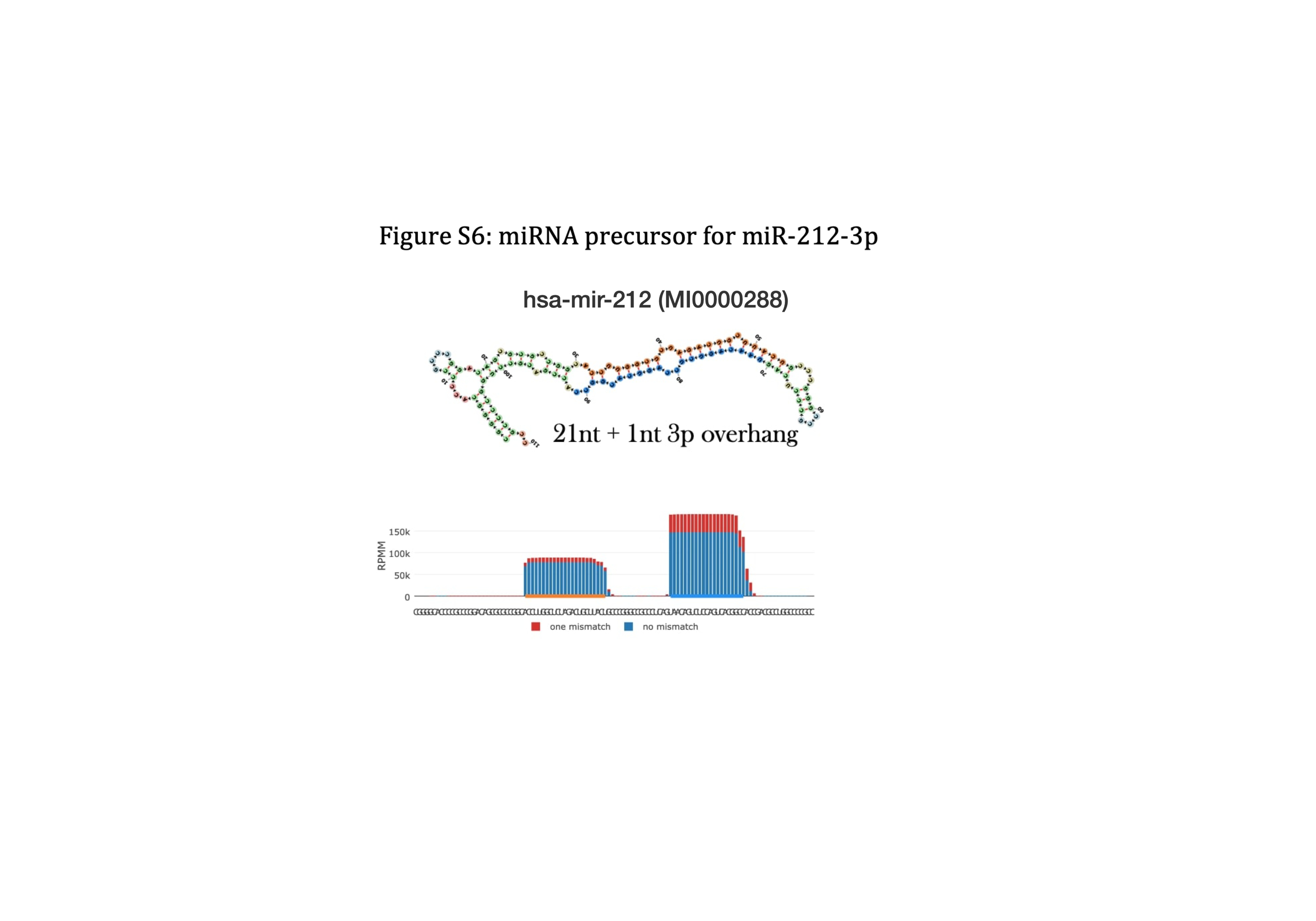
miR-212-3p has previously been linked to treatment of depression. In rats subjected to electroconvulsive stimulation, the animal model equivalent to electroconvulsive therapy, regulation of 6 miRNAs in the HIP has been reported, among them upregulation of miR-212-3p31. The 5p transcript (miR-212-5p) has also multiple times in the literature been linked to depression. According to the sequencing density in miRCarta and miRbase, miR-212-3p appear to be the high-abundance/guide strand in humans (Figure S6). Therefore, miR-212-5p has not previously been annotated, and thus, not included in the Nanostring v3b panel by the manufacturer. Hence, miR-212-5p could not be quantified in the pigs. In depressed patients, miR-212 has been reported to be elevated in serum after treatment with selective serotonin re-uptake inhibitors (SSRIs) and serotonin and norepinephrine re-uptake inhibitors. It is unclear whether it was the 3p or the 5p strand, that was investigated32. To further explore the possible link between depression and miR-212, the antidepressive potential of miR-212 was investigated in a chronic unpredictable mild stress (CUMS) mouse model of depression (Si et al. (202133)). Although not stated explicitly, it was indicated that miR-212-5p was the focus of the study. The CUMS mice exhibited various depressive-like behaviours, determined by body weight, sucrose preference test, forced swimming test, and tail suspension test which all could be ameliorated by either fluoxetine or over-expression of miR-212-5p in the HIP33. In addition, it was demonstrated with Targetscan predictions and in luciferase assays, that miR-212-5p targets and downregulates nuclear factor IA (NFIA) and that the antidepressive-like effects mediated by miR-212-5p were abolished when combined with NFIA over-expression33. NFIA regulation may thus be a necessary mechanism underlying the antidepressant-like effects of miR-212-5p. In the context of bladder cancer, it has been demonstrated with Targetscan predictions and luciferase assays that miR-212-3p also targets NFIA34, thus demonstrating that both miR-212-5p and miR-212-3p targets NFIA and may act through similar antidepressive molecular mechanisms. Interestingly, in a study by Si et al. (2021), the depressive-like CUMS mice already exhibited elevated miR-212-5p levels in blood and HIP, but the antidepressant like effects were achieved through further over-expression of miR-212-5p33. This indicate that upregulation of miR-212-5p might be a protective compensatory mechanism in CUMS mice, rather than a pathological mechanism that needs to be corrected. In support of this assumption, Si et al. (2021) provided evidence for neuroprotective and anti-inflammatory (reduced Tumor necrosis factor-alpha (TNF-α), Interleukin-1 beta, and Interleukin-6 levels) effects using miR-212-5p mimics in the CUMS mice33. Similarly, psilocybin has been reported to acutely decrease TNF-α levels in blood from healthy subjects, as TNF-α was reverted to baseline levels, one week after psilocybin administration35. This timeline is comparable to the duration of the miR-212-3p regulation observed in the pigs in our study – upregulated after 1 day, normalized after 7 days. In addition, it has been reported that psilocybin decreased TNF-α levels in human U937 macrophage cells36. This could indicate that part of the underlying antidepressive mechanism of psilocybin is to acutely induce neuroprotective and anti-inflammatory effect through expression of miR-212 which acts on NFIA, which in turn regulate TNF-α levels. These results align with prior RNAseq studies performed on the same pigs, which demonstrated that the vast majority of regulated pathways, were immune-related11.
Lastly, miR-212 has also been observed down-regulated in monocytes from patients with post-partem psychosis (generally thought to belong to the bipolar spectrum)37 and downregulated in post-mortem tissue from patients with bipolar disorder38.
miR-107 is downregulated 1 day after psilocybin administration in PFC
miR-107 and miR-103-3p are sequence homologs and likely exhibit the same biological functions (Auwera39 and miRbase.org). Upregulation of miR-107 has been observed in whole blood from manic bipolar patients compared to healthy controls40. On the contrary downregulation of both miR-107 and miR-103 has been reported in plasma from psychiatric patients with childhood trauma39, which is one of the best predictors of depression later in life41. Transforming growth factor beta receptor 3 (TGFBR3) was identified as a target for both miRNAs39. In parallel, SNPs which associated with response to antidepressive treatment in a pool of 575 depressed patients were reported42. The rs12082710 genotype (SNP within the TGFBR3 gene) achieved the best association and from a total of 14 identified SNPs which associated with antidepressive treatment outcome, 9 were located within TGFBR342. TGFBR3 encode betaglycan, a co-receptor mediating functional antagonism of activin signalling. Hence, it has been demonstrated that injection of Activin A into the mouse HIP provides an antidepressant-like effect42. In resemblance with NFIA, Activin has been shown to provide neuroprotective and neuroplastic effects43. While the precise nature of the relationship between the pathophysiology of depression and neuroplasticity is complex, it is generally accepted that antidepressants also interfere with neurotrophic factors which modulate neuroprotective and neuroplastic effects44–48. It is therefore indicated that miR-107 and its target TGFBR3 might play a key role in depression and antidepressant treatment. However, this might be an oversimplification as our data showed downregulation of miR-107 in the pigs 1 day after psilocybin treatment, which indicate upregulation of TGFBR3, which provide functional antagonism to the antidepressant effects of activin. However, according to Targetscan, miR-107 has 907 conserved target sites across 825 transcripts and might therefore exhibit an array of different functions. In support of this statement, it has been demonstrated that gastrodin ameliorates the neuroinflammatory effects induced by lipopolysaccharide in mice, and that the functional mechanism rely on gastrodin-induced downregulation of miR-107 (overexpression of miR-107 abolish the anti-inflammatory effects of gastrodin)49. The observed downregulation of miR-107 in the pigs might thus indicate anti-inflammatory effects in resemblance with the effect of miR-212 overexpression33.
Overall, it seems that miR-107 upregulation can provide antidepressant effects through reduced TGFBR3-mediated activin antagonism, while miR-107 downregulation can play into the gastrodin pathway causing anti-inflammatory effects. This dualistic effect might explain the diverging observations in psychiatric patients with childhood trauma39 and bipolar patients40. Nonetheless, the observations in patients were based on peripheral blood samples, which might not correlate with regulations in the brain.
miR-98-5p and miR-128-3p are downregulated 1 day after psilocybin in PFC
It has previously been reported, that among 25 regulated miRNAs, miR-128-3p was upregulated in the amygdala from the learned helplessness rat model of depression50. Prediction target analysis revealed genes within the Wnt signalling pathway as miR-128-3p targets, and significantly reduced Wnt signalling genes in the amygdala of the depressive-like rats was observed50. In parallel, elevated miR-128-3p levels and decreased Wnt signalling genes (WNT5B, DVL, and LEF1) were also observed in post-mortem amygdala from depressed patients50. This is in line with other studies during recent years, as accumulating evidence implicate Wnt signalling in neuropsychiatric disorders, neurogenesis, and in the underlying mechanism of mood-stabilizers51, 52. In mice subjected to chronic restraint stress (CRS) reduced levels of Wnt2 and Wnt3 in the ventral HIP have been reported53. Furthermore, knockdown of Wnt2 and Wnt3 led to impaired Wnt/β-catenin signalling, neurogenesis deficits, and depression-like behaviours53. In contrast, overexpression of Wnt2 or Wnt3 reversed depression-like behaviours in the CRS mice53. Lastly, treatment with the selective serotonin re-uptake inhibitors fluoxetine increased Wnt2 and Wnt3 levels in the ventral HIP and Wnt2 or Wnt3 knockdown abolished the effect of fluoxetine53. Overall, the results presented here, indicate that when psilocybin is given to pigs Wnt signalling is increased through decreased miR-128-3p levels which in turn might mediate antidepressive effects.
In contrast to our results, it has been reported that miR-98-5p is downregulated in the PFC and HIP from mice subjected to chronic social defeat stress and that miR-98-5p overexpression alleviated the depressive-like behaviours54. Furthermore, 1 day after ketamine administration, miR-98-5p was elevated and miR-98-5p inhibition blocked the antidepressant effect of ketamine54. In our study, miR-98-5p was the only miRNA with the expression levels on day 1 and day 7 being in opposite directions, a significant regulation was observed between PSI-1-day and PSI-7-day (Fig. 1B). Thus, it can be speculated that there is a time-delayed effect of psilocybin on miR-98-5p, and a greater magnitude of effect might be observed by using stressed/depressive-like pigs.
miR-92a-3p and miR-485-3p are downregulated 1 day after psilocybin in HIP
Dysregulated miR-92a-3p has previously been linked to depression in clinical studies. It has been reported that miR-92a-3p is downregulated in the dorsolateral PFC from suicide subjects and this was inversely correlated with TNA-α levels, despite not being a direct target of miR-92a-3p55. In contrast, subjects with high baseline miR-92a-3p are more likely to develop post-stroke depression56. Preclinically, antidepressant effects has been coupled with reduced miR-92a-3p levels in the HIP of CUMS rats and inhibited by AAV-mediated over-expression of miR-92a-3p57. In the CUMS rats the depressive-like behaviour was ameliorated, when the rats were exposed to enriched environment (EE). EE reduced neuronal apoptosis and increased Tropomyosin receptor kinase B (TrkB) and Brain-derived neurotrophic factor (BDNF) levels in the rats, this was partially reversed by miR-92a-3p over-expression57. Reduction in miR-92a-3p might thus provide a neuroprotective and neurotrophic effect in resemblance with conventional antidepressants. miR-92a-3p was one among only two miRNA that was regulated (down-regulated) in the HIP of the pigs after 1 day, and it remained approximately significantly regulated in the late phase (p = 0.0509, Fig. 2). This could indicate that psilocybin induces a neuroprotective and neurotrophic effect through downregulation of miR-92a-3p. To further support this statement, Muñoz-Llanos et. al. reported that miR-92a-3p was elevated in the dorsal HIP of male rats exposed to 14 days of restraint stress. They also reported that miR-485-5p (although not miR-485-3p) was upregulated in the HIP of the restraint stress rats30. In silico analysis revealed that miR-92a-3p and miR-485-5p share many biological functions30. miR-485-5p was expressed below LOD in the HIP of the pigs and could thus not be reliably detected. Conversely, miR-485-3p has been reported upregulated in peripheral blood mononuclear cells (PBMCs) of depressed patients after 8 weeks of administration of various antidepressants (week 0 versus week 8)58. However, regulation in peripheral blood might not reflect changes in the HIP and the use of various antidepressants in the study, might not exert the same regulatory effects as psilocybin.
miR-99b-5p, miR-125a-5p, and miR-221-3p are downregulated 7 days after psilocybin in HIP
As none of the three miRNAs were regulated after 1 day (Fig. 2A), this could suggest the presence of distinct antidepressant regulatory mechanisms operating at different time points following psilocybin treatment.
Previously, miR-99b-3p (but not the guide strand - miR-99b-5p) has been reported upregulated in post-mortem brains of suicide subjects59 and plasma miR-99b (strand not specified, but we assume detection of guide – miR-99b-5p) is downregulated after twelve weeks of escitalopram treatment in depressed individuals60. Similarly, miR-221-3p is upregulated in serum from depressed subjects61 and miR-125a-5p has been reported to be upregulated in plasma and cerebrospinal spinal fluid from depressed individuals62, 63. Likewise, miR-125a-5p was upregulated in PFC of mice subjected to acute and repeated stress64, but downregulated in the HIP of CUMS rats65.
Thus, the psilocybin-induced downregulation of miR-99b-5p, miR-125a-5p, and miR-221-3p appear to counteract the general upregulations observed in depressed patients and depressive-like animals.
Using lentiviral plasmid overexpression of miR-221-3p in HA1800 cells, it has been demonstrated that miR-221-3p target and downregulate interferon regulatory factor 2 (IRF2), a negative regulator of interferon-alpha (IFN-α)61. IFN-α is a cytokine closely associated with depression66 and it is thus suggested that the psilocybin-induced downregulation of miR-221-3p lead to reduced inflammation by downregulating IFN-α through upregulation of IRF2.
In conclusion, we have demonstrated that the Nanostring nCounter technology and the predefined Human v3b miRNA Assay panel can be used to explore the miRNA landscape in pig brain tissue. Further, we present evidence that psilocybin may exert its intracellular antidepressant effects through miRNA mechanisms. miR-92a-3p, miR-98-5p, miR-99b-5p, miR-107, miR-125a-5p, miR-128-3p, miR-212-3p, and miR-221-3p have been identified as specific miRNA targets. Previously, these miRNAs have been proven to exhibit antidepressant potential in animal models of depression, however, none of them have been associated with psychedelic effects. The combined effect of these miRNAs might therefore exert the antidepressant potential of psilocybin without aversive psychedelic effects. Further studies are needed to elucidate both the acute and prolonged effect of the psilocybin regulated miRNAs and their downstream targets.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}