Plant lectins are usually found in storage organs such as seeds 8, 21, however, could also be found in leaves, stems, cortex and fruits, varying in concentration according to the plant organ evaluated4,22.
There are very few reports referring to the extraction of plant lectins from vegetative tissues, most of them based on the use of saline solutions, some of them present variations on the pH, or consider the addition of other compounds, such as ascorbic acid, Triton X-100, among others13,21,22.
Comparing the yield reported by Adenike y Babalola in 200423 for Kalonche crenata leaves, Yan et al in 200924 for Astragalus mongholicus roots and Liu et al. in 201025 for Pinellia pedatisecda stems whom obtained a total protein yield of 4.8%, 1,8% y 3,5%, respectively, which is similar to the yield obtained in this study, even though the extraction method used was PBS buffer, having a medium ionic force., while Andrade et al. in 200526, for Bauhinia monandra leaves, and He et al. in 201127 for Ophioglossum pedunculosum roots obtained a 1.6% proteic yield, using a NaCl solution, confirming that saline solutions are an adequate method of total protein extraction, allowing to obtain plant lectins.
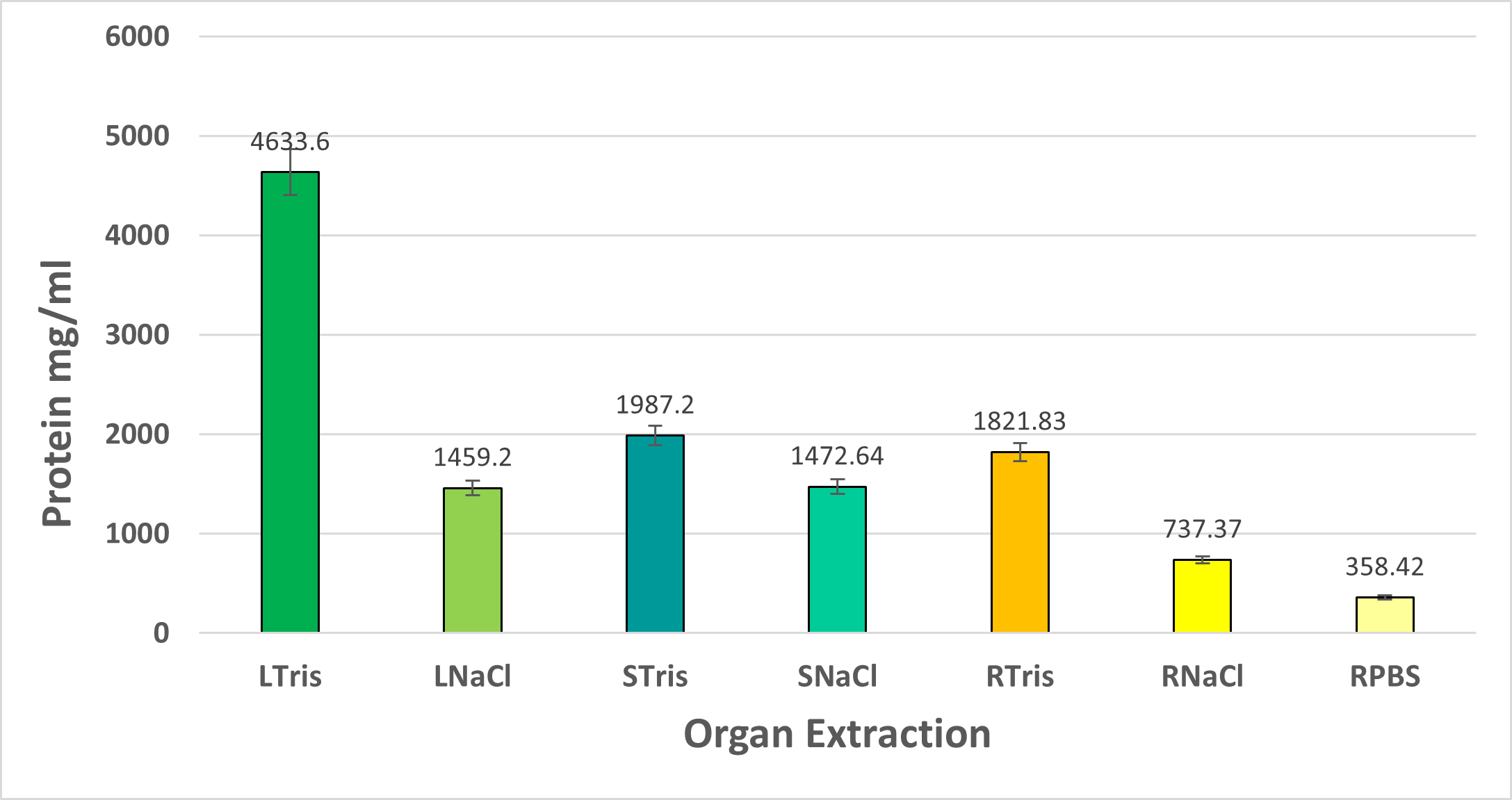
In this study two plant lectin extraction methods were assessed, obtaining through the NaCl extraction method a yield of total protein of 1.45% in leaves, 1.47% in stems and 0.737% in roots, while using Tris-HCl extraction method, the protein yield was 4.63% in leaves, 1.98% in stems, and 1.82% in roots.
According to these results, Tris-HCl extraction methos allows to obtain a bigger protein concentration, confirming that this method is adequate to obtain a bigger amount of total protein.
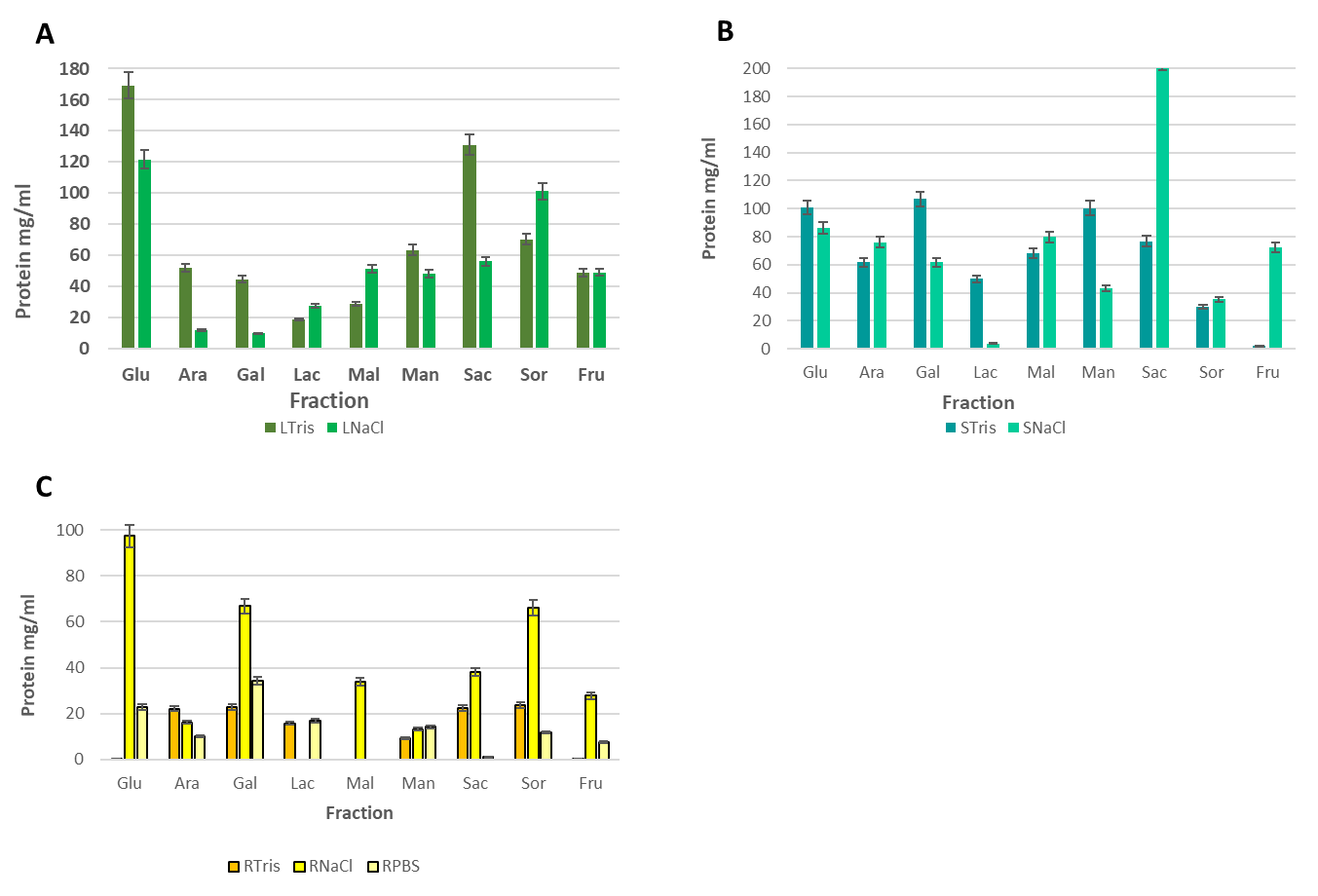
After demi purification methods were performed, through dialysis and affinity chromatography, lectin-like proteins yield, comparing to total protein yield, was 32.5% in leaves extract, 45.3% in stem extract and 48.78% in root extract, through the NaCl extraction method, while using the Tris-HCl method, the lectin-like protein yield, comparing to total protein obtained was 13.49% in leaves extract, 30% in stem extracts and 6.58% in root extract; opposing to what Torres reported in 200628, who stablished the Tris-HCl extraction method as the best to obtain lectin-like proteins, as our results supports that a bigger yield can be obtained through a high- ionic force solution, such as NaCl.
In the other hand, our results differ from the ones reported by Flores in 20122, where lectin-like protein obtained from Ruta graveolens yields through the NaCl extraction method were 3.34%, 2.55% y 1.51% out of leaves, stem, and roots, respectively. These results could be related to seasonal differences in different molecules yielding, as this study was performed near blooming season, while 2012 study was performed through January and February. There’s evidence sustaining seasonal changes in lectin production trough blooming and Winter seasons22. (22).
A widely reported lectin-like protein demi purification technique was affinity chromatography, which exploits the high affinity to certain chemical groups establishing non-covalent bonds, and then being eluted using a free ligand29,30.
The most commonly used matrixes for lectin-like protein purification are Sephadex, which is a glucose complex with free OH y CH2 groups, and Sepharose, which consists of two galactose molecules with free CH2 y OH groups, which could also have active thiol, oxirane and ester groups, allowing binding to C = O, N = N and C = C structures, interacting with OH, NH2 y SH groups, however, acrylamide and silica gel matrixes have also been reported in this context31.
Silica gel has two oxygen molecules bound to silica trough double bounds, getting a similar structure to the one found in an ester group, allowing the protein and silica gel interactions and connections. Pereira et al. in 201513 reported the use of a silica gel matrix as a second demi purification step to obtain Colocasia esculenta lectins, while in this study a silica gel matrix was used in a simple chromatography, where lectin-like proteins were obtained.
Other studies stablish glucose elution as the main method to separate lectin-like proteins from chromatography columns5, 7,32, however, the yield reported by Flores in 2012 allowed to obtain from 6 to 13% of lectin-like proteins from Ruta graveolens, while this study’s results stablish a yield ranging from 32 to 49%, which can be attributed to the use of 9 different eluents in the affinity chromatography, allowing to consider the existence of a wider variety of lectin-like proteins, with affinity to different carbohydrates.
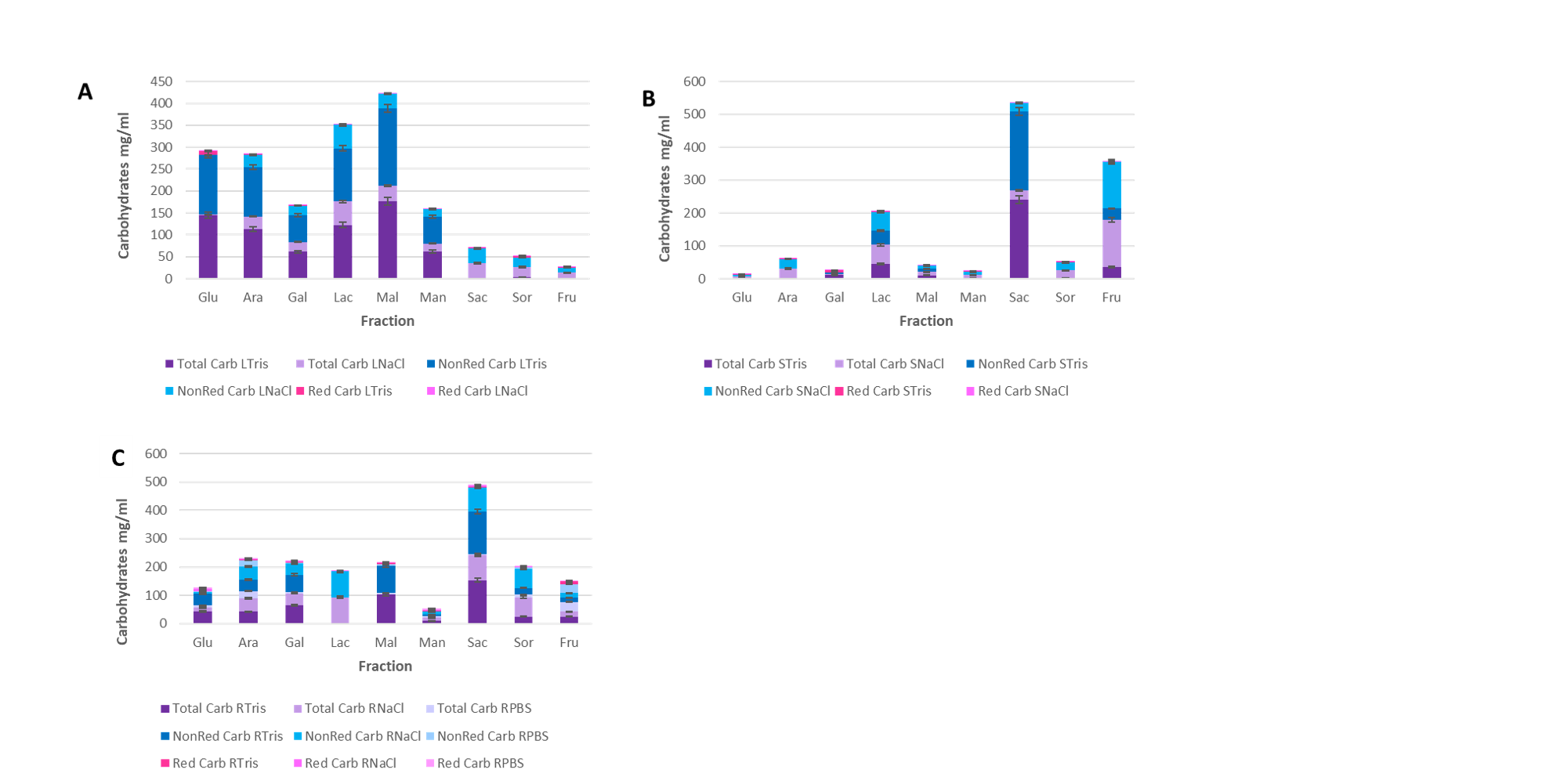
Total sugars, reducing and non-reducing sugars were quantified as part of the partial characterization of the different fractions, where data suggest that there is a bigger abundance of non-reducing sugars in the protein structure, adding up to 95% of all the sugars in fractions obtained from R. graveolens, with a protein-carbohydrate ratio of 0.1 to 5 mg, however, most carbohydrates associated with lectin-like protein structures reported are reducing sugars12 (18), which constitutes oligosaccharide elongation cores, while non-reducing sugars could be part of the peripheral region.
Furthermore, comparing both extraction methods, even though Tris-HCl extracted fractions had a bigger protein concentration, had a smaller protein-carbohydrate ratio, suggesting that the lower ionic-force method could allow to obtain different lectin-like proteins, against the possible obtainable lectin-like proteins through NaCl extraction methods. Another possible explanation could be the loss of the glycosidic bond when diminishing the carbohydrate concentration.
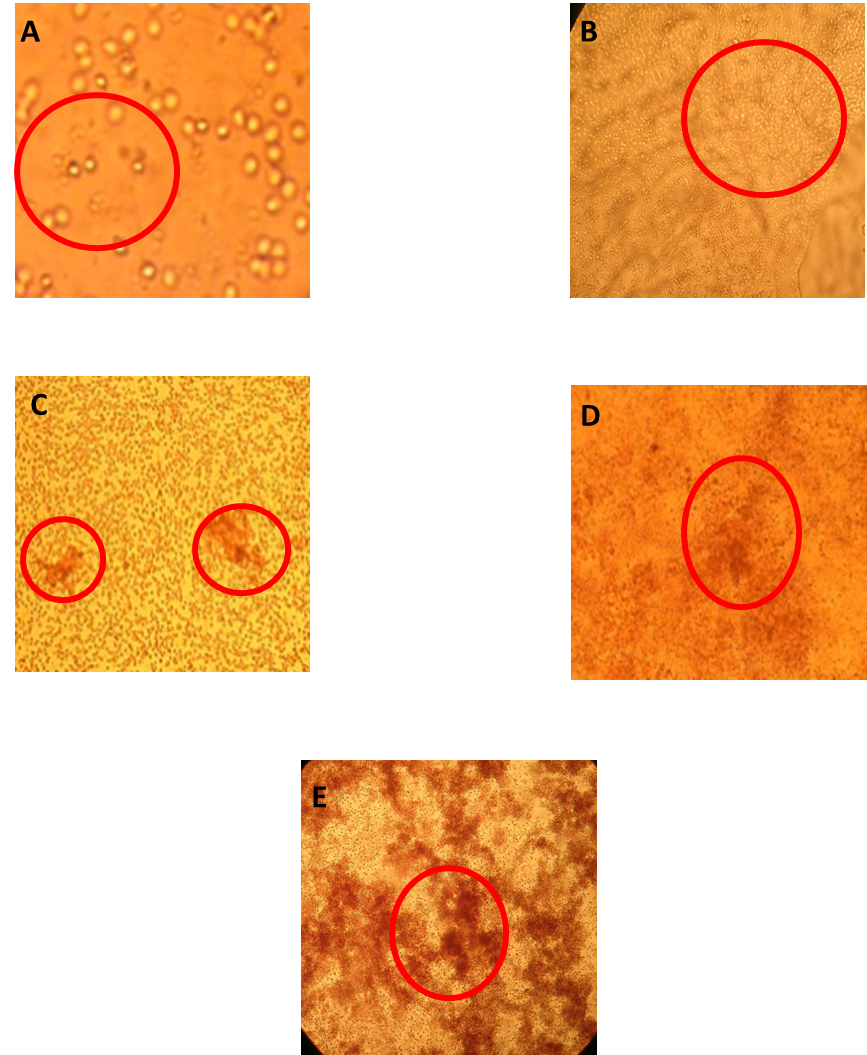
Lectin-carbohydrate interactions are assessed through the hemagglutination assay, process, getting two crossing types: I) a bivalent lectin-like protein crossing with a monovalent carbohydrate in one dimension and II) multivalent lectins and carbohydrates crossing, as one of the molecules has a bigger valence and creates two or three dimensions complexes that could precipitate when in solution33, being the latter the crossing experienced in hemagglutination processes.
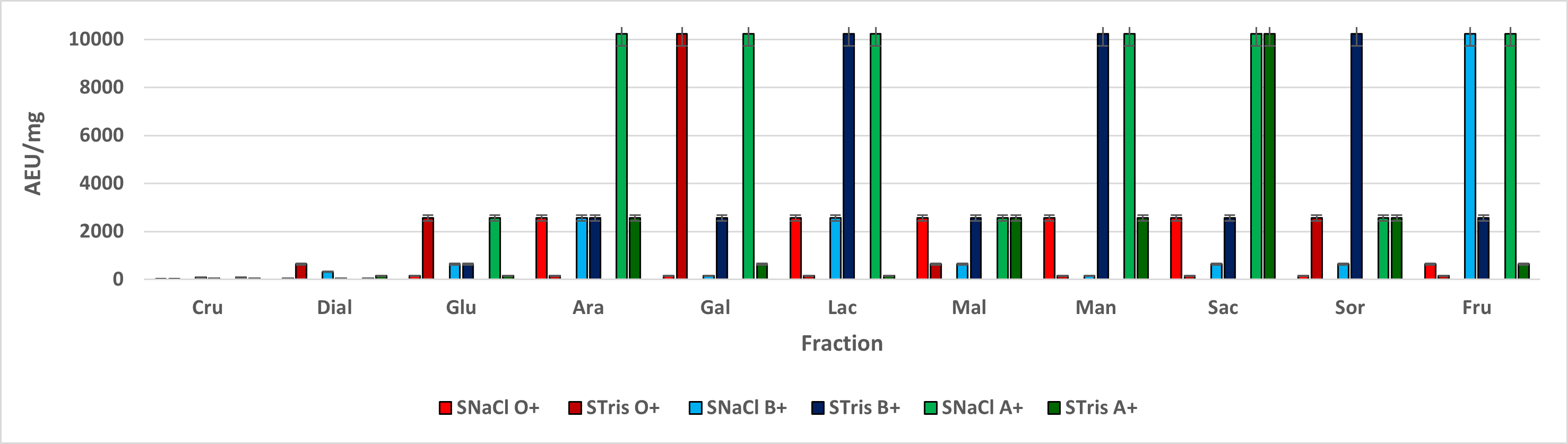
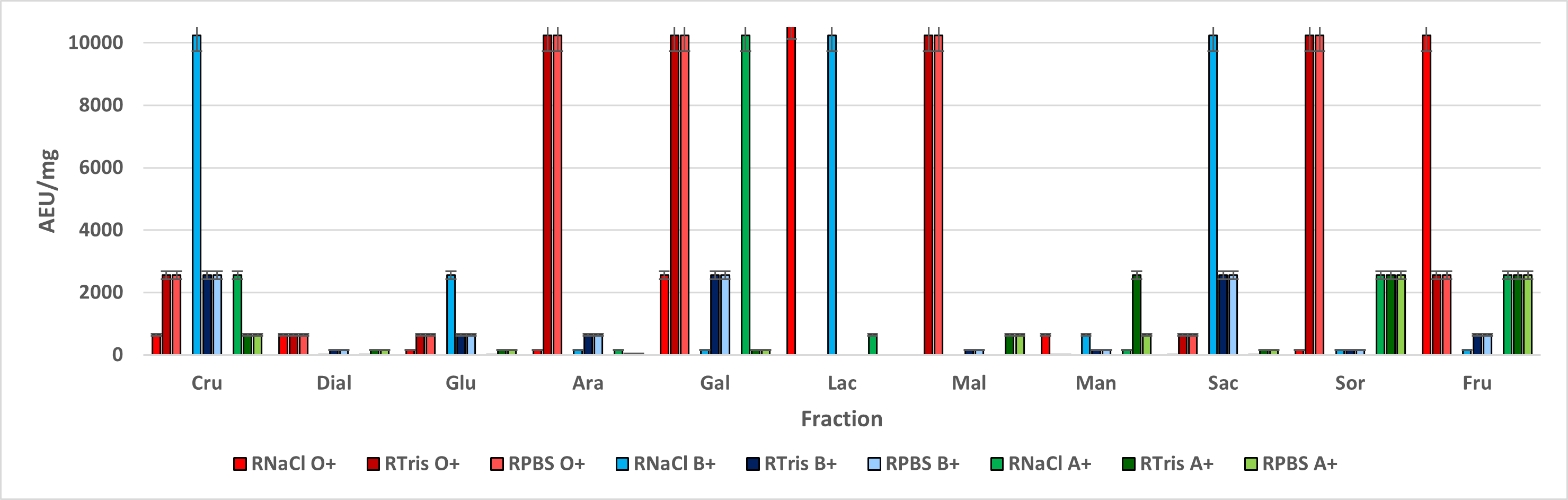
The bigger hemagglutination effect was assessed when fractions from leavers and stem interacted with A + blood type, while root fractions had a bigger effect when interacting with root fractions. These results agree with the ones obtained by Hernández in 20111 and Flores in 20122, suggesting a higher affinity to N-acetyl galactosamine from leaf and stem lectin-like proteins, while root lectin-like proteins had a higher affinity to L-fucose.
Even though R. graveolens lectin-like proteins seem to have higher affinity to certain blood type, hemagglutination activity in other blood type red cells was assessed, so it’s possible to classify them as non-specific lectin like proteins23. However, it’s important to consider that fractions were demi purified, so it’s possible that we could have an unspecific mix of lectin-like proteins, or the presence of more than one recognition domain.
It’s important to consider that temperature, pH, and metallic ion binding could affect the non-covalent interactions present in non-covalent interactions, leading to total or partial loss of lectin-like protein structure and biological activity 34,35.
Lectins are amphoteric and, as such, they can accept or donate protons, so pH changes affect their ionization state, changing its isoelectric point, resulting in Van Der Waals forces changes, promoting structure changes29,30 thus, metallic bonds and their biological effect can be affected. This could explain the differences among fractions obtained from the same tissues, eluted with the same sugar but extracted with a different saline solution, being NaCl extracted fractions the ones with a higher AEU/mg range, comparing to the ones extracted with Tris-HCl buffer.
According to Hernández in 20111 and Torres in 201536, the pH stability range of R. graveolens lectin-like proteins is ranging from 5.4 to 8.4, even though these differences could be associated to pH differences in both solutions, both have a pH located in the stability range, suggesting different lectin-like proteins as the main possible reason why the hemagglutination effect is different. Loss of structural carbohydrates, ionization changes or metallic ion sequestering by Tris-HCl could be cited as possible structural change causes, thus changing the affinity and exposition of specific binding sites20 .
Comparing both extraction methods through hemagglutination kinetics, it was observed that similar protein concentrations led to similar agglutination levels, suggesting that this activity relates to the concentration needed to allow the agglutination to happen, while agglutination level relates to the number of molecules that could bind to a cell or lectins.
Furthermore, comparing among the same agglutination level simples, incubation time was higher for those extracted with low-ionic force solutions, such as Tris-HCl.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}