We conducted an in-depth, large population-based analysis of the effect of yoghurt consumption on the gut microbiome characterised via 16S rRNA and whole shotgun metagenomic sequencing while accounting for covariates, such as age, gender, BMI and most importantly habitual diet. Analysis of 16S rRNA gene sequencing data but not whole shotgun metagenomic sequencing on a lower sample size, revealed that gut microbiota from yoghurt consumers harboured a higher alpha-diversity than that of non-consumers. In an independent cohort (LifeLines-DEEP), shotgun sequenced data and 16S rRNA, alpha diversity was not increased in the gut of yoghurt consumers, which may be due to the fact that only a low proportion of the population (8.4%) could be clarified as non-yoghurt consumers, limiting statistical power. Taken together, our results suggest that larger sample size and comparable group size is needed to elucidate the contribution of yogurt consumption to gut microbiota alpha diversity, obscured by technical and population variability.
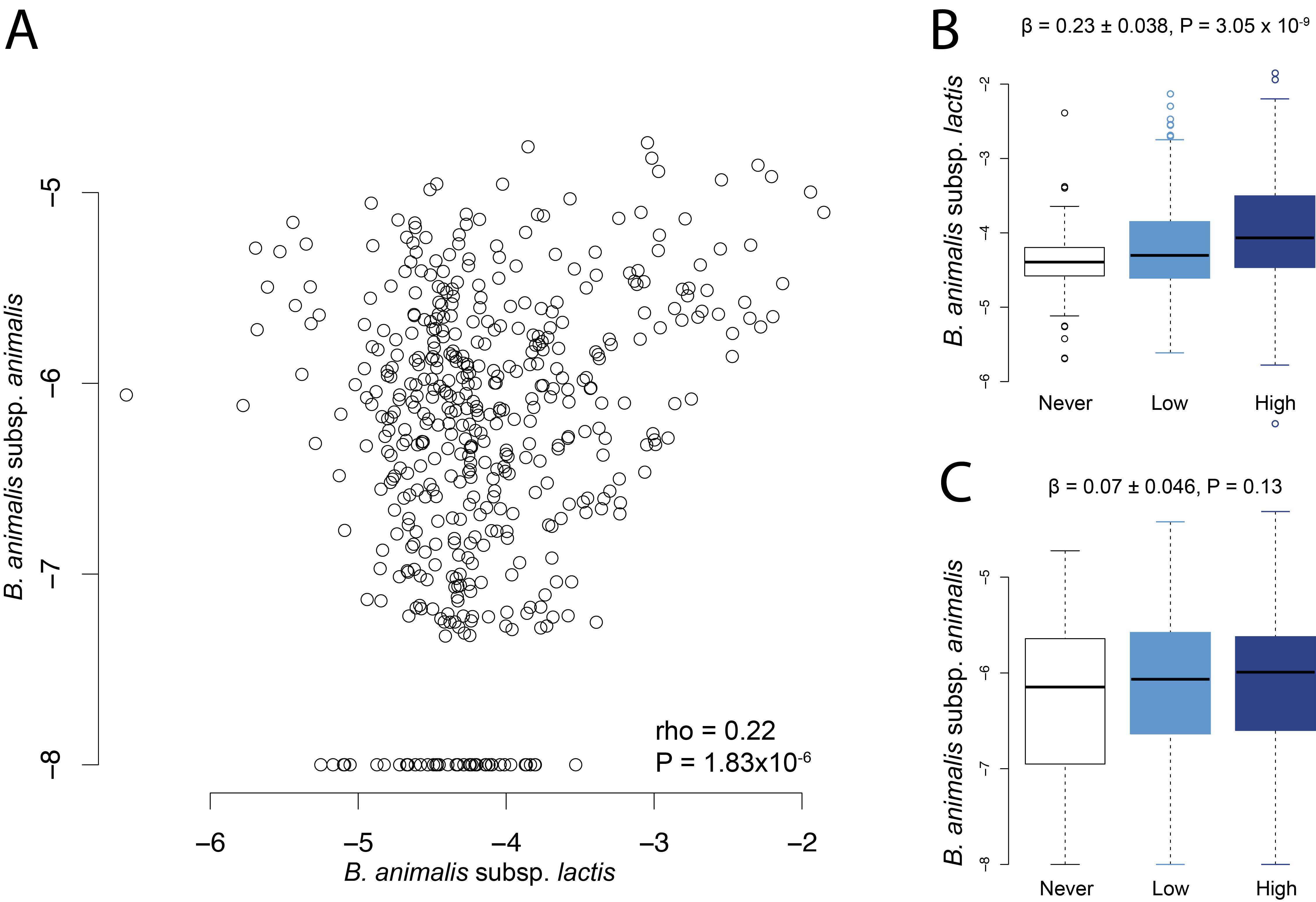
Yoghurt consumption was associated with an increase in Streptococcus but also Christensenella and Ruminococcaceae. The observed increase in Ruminococcus genus in our study is in contradiction with previously published results where a 24-week yoghurt intervention was followed by a decrease in Ruminococcus [6]. This discrepancy may be due to the differences in population and statistical analysis. We observed an increase in Streptococcus with 16S RNA gene sequencing that was further assigned to S. thermophilus by Shotgun sequencing, also observed in LifeLines-DEEP cohort. Besides, B. animalis subsp lactis added to yoghurt products, was found to be increased in the yoghurt consumer group of the TwinsUK cohort. Both bacterial species/subspecies are used in the making of fermented milk products and were found to be increased in the gut of yoghurt consumers in several other observational studies [19, 20]. Yet, the B. animalis subsp lactis observation was not replicated in the LifeLines-DEEP cohort, potentially indicating that dairy products consumed by participants of in the LifeLines-DEEP cohort were less enriched in B. animalis subsp lactis. Since B. animalis subsp lactis is a species adapted to the GI tract and known to reach the colon alive, one can make the hypothesis that the higher alpha diversity observed in the TwinsUK is specific to the consumption of B. animalis subsp lactis enriched products, less consumed in the the LifeLines-DEEP cohort. Thus, following yoghurt consumption, gut microbiome composition is characterised by an increase of the bacteria ingested from the product, which also implies that a fine description of the bacteria composition of yogurt is needed to further define their effects on the gut microbiome.
Using a combination of FFQs and food record data we observed that the increase of these two bacteria in the human gut following yoghurt consumption might be transient as highly determined by consumption of yoghurt within the 24 h preceding the faecal sample. Transient detection of yogurt / fermented milk strains has been shown in various clinical studies [13, 39]. While S. thermophilus and B.animalis subsp lactis are able to thrive in simple communities, they are outcompeted by resident bacteria in more complex ecosystems through colonisation resistance. Our data confirm that yoghurt-bacteria are transient members of the gut microbiome and do not durably engraft within the gut lumen. Interestingly, a recent study showed that Streptococcus thermophilus was one of the most prevalent lactic acid bacteria detected in 9445 metagenomes from human samples [40].
Co-occurrence network analysis indicated that B. animalis subsp lactis and S. thermophilus belonged to a subnetwork composed exclusively of lactic acid bacteria, with a co-variance most likely explained by yoghurt consumption. This may relate to heterogeneity in the permissiveness of the resident ecosystem to integrate ingested strain [41–45]. It is not possible to exclude that both bacteria, B. animalis subsp. lactis and S. thermophilus, may exert a greater impact on the microbial ecosystem in the small intestine where ingested strains outnumber resident microbes [17]. Alternatively, yoghurt bacteria might exert their effect locally as they can adhere to some extend to intestinal cells [12], which would need to be ascertained using more invasive technics [41]. Taken together, our study shows that while cross-sectional cohort can reveal association between transient microbes and gut resident gut microbiome, longitudinal settings coupled with population stratification and alternatively biogeographical sampling of the gut microbiome are warranted to better decipher the detailed nature of these interactions. Besides, these results infer that the design of fermented dairy product may in the future benefit from the addition of strains capable of integrating and / or covarying with the gut ecosystem more efficiently in the context of precision medicine [46].
We observed that B. animalis subsp. lactis was associated with 13 faecal metabolites. Among these, we reported a positive association with 3-hydroxyoctanoic acid, an agonist of the hydroxycarboxylic acid receptor 3 (HCA3) that was not reflected in blood. HCA3 is expressed in enterocytes and its inactivation by 3-hydroxyoctanoic acid mediates a reduction of inflammation [47]. Our results are in line with a recent study demonstrating that metabolites of lactic acid bacteria present in fermented foods, i.e. 3-hydroxyoctanoic acid can inactivate HCA3 [48]. In accordance with the literature, we reported that yoghurt consumption was associated with markers of metabolic health, namely a decrease in VFM and fasting insulin. Even though these two parameters have previously been linked with gut microbiome alpha-diversity and the abundance of some genera such as Christensenella, that were both increased in the gut of yoghurt consumers [49–51], neither of the two yoghurt bacterial species were linked to these phenotypes. This implies that factors others than the increase in S. thermophilus and B. animalis subsp. lactis may also be at play.
As previously described in epidemiological studies [52–56], we reported that yoghurt consumption was linked with healthy dietary habits. In the present study we intended to account for this bias by adjusting for habitual diet which in part attenuated the original results. However, the HEI used here as a covariate may not capture in full diet quality and specifically, the individual contribution of fruit, whole grain and protein that were significantly different between yoghurt consumers and non-consumers. The HEI was generated based on FFQs that may be biased by the fact that volunteers tend to over report healthy foods in self-reported questionnaires [57]. The latter may also have impacted the assessment of yoghurt consumption. Further, one of the main limitations of the present work is its inherent observational nature that does not allow inference of causal relationships. Finally, associations reported here were observed in a predominantly older white female British cohort and may not apply to other populations. Nevertheless, some of the microbiome results were replicated in an independent Dutch cohort and generally reflected current literature suggesting observations reflect wider patterns applicable to other populations.
{kind=link}