3.1 Elicitation
3.1.1 Effects of elicitor type and concentration
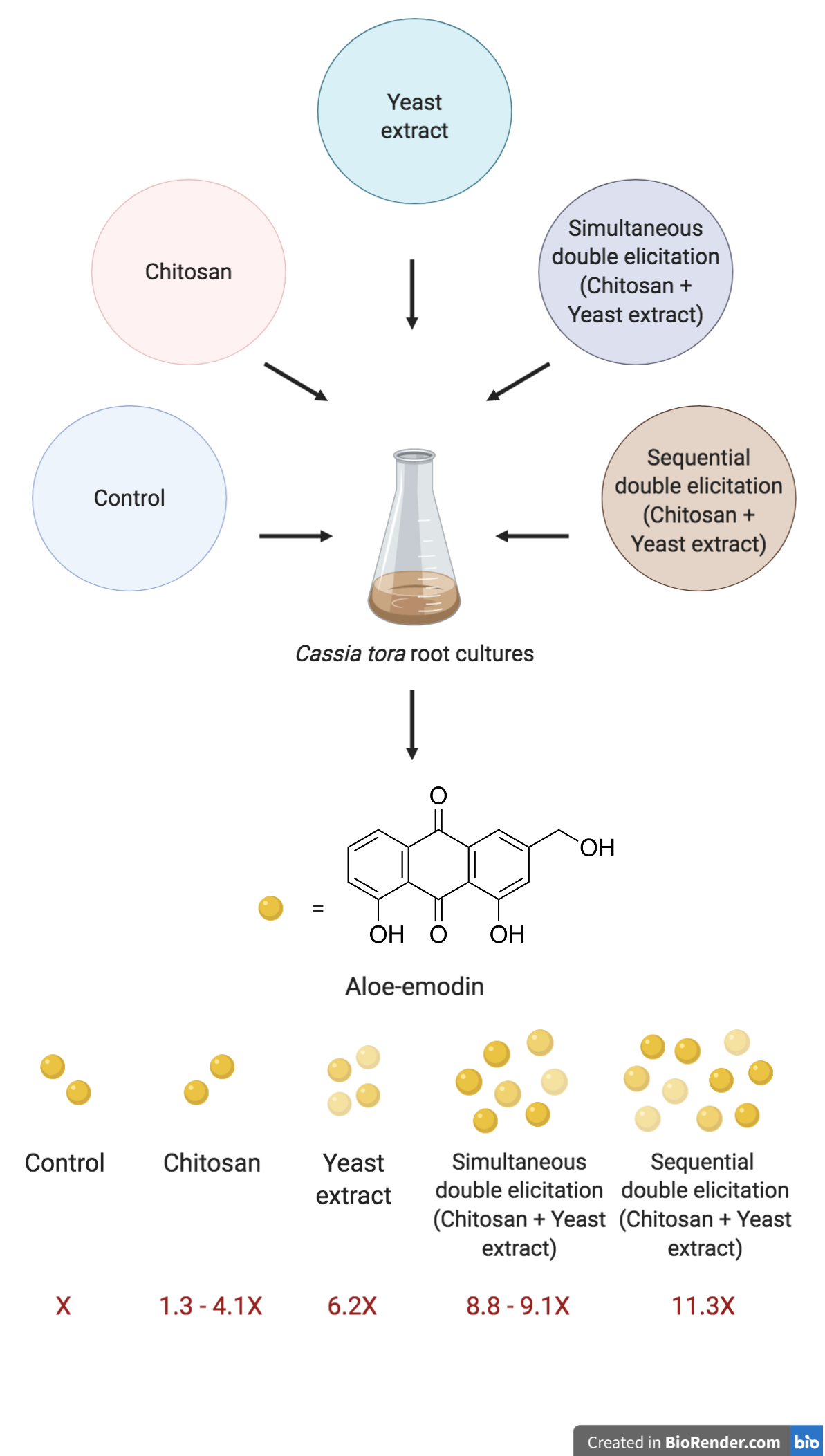
Chitosan and yeast extract have been frequently reported as successful elicitors for enhancing anthraquinone production in many plants, such as C. angustifolia (Chetri SK et al., 2015), C. bicapsularis (Abdel-Rahman IAM et al., 2013), Rudgea jasminoides (Oliveira MDC et al., 2007), Oldenlandia umbellate (Krishnan RS and Siril EA, 2016), Rubia akane (Jin JH et al., 1999), R. tinctorum (Vascosuelo A et al., 2003) and Myristica fragrans (Iyer I et al., 2011). In this study, the un-elicited root cultures began the production of aloe-emodin on day 12 of the cycle and reached the peak level on day 20. The varied concentrations of yeast extract and chitosan were used to enhance the production. The elicitation period was initially set at 24 hours to avoid cytotoxicity caused by long-time contact with the elicitor. The elicitation effect was found to be highly influenced by the type and concentration of elicitor. At 10 mg/L, both elicitors failed to enhance the production. However, their effects gradually improved as the concentration increased. At 50 mg/L of chitosan, the production level was elevated, but still not better than the control. In comparison, 50 mg/L of yeast extract significantly increased the production by 1.68 times when compared with the control (0.391 ± 0.024 mg/gDW). The peak performance of chitosan was observed at the concentration of 100 mg/L. It increased aloe-emodin production from the control by 1.18 times (0.276 ± 0.007 mg/gDW). While, the peak effect of yeast extract was also observed at 100 mg/L but it yielded a higher level of aloe-emodin than chitosan did (Fig. 1). Yeast extract at 100 mg/L was the most effective elicitor, increasing aloe-emodin production up to 2.71 times (0.633 ± 0.048 mg/gDW) when compared with the control. Nevertheless, the concentration-dependent manner only applies to the particular range of concentrations. At relatively high concentrations, the effects of chitosan and yeast extract were dropped from their peak level. Furthermore, these elicitors differently affected the accumulation in both intracellular and extracellular compartments. Yeast extract promoted a comparable accumulation in both compartments, whereas chitosan had a distinct advantage toward the intracellular accumulation (Fig. 1).
The results suggested that C. tora root cultures were more susceptible to yeast extract than to chitosan. Interestingly, similar findings were also observed in C. angustifolia (Chetri SK et al., 2015) and C. bicapsularis (Abdel-Rahman IAM et al., 2013). Yeast extract showed potential to enhance Cassia sp. anthraquinone production, as well as other acetate-malonate-derived anthraquinones (Krishnan RS and Siril EA, 2016; Oliveira MDC et al., 2007). Meanwhile, chitosan was reportedly efficient in Rubiaceae (Vascosuelo A., et al., 2003; Jin JH et al., 1999) and other species producing Rubia-typed anthraquinones (Iyer I et al., 2011). According to current knowledge on the plant defence system, it might be assumed that the different preference for chitosan and yeast extract is contributed by the different type and number of defence components expressed in the two genera.
Normally, plants respond to pathogen stress through the induction of pattern-triggered immunity (PTI). The epitope part of pathogen or the so-called pathogen-associated molecular pattern (PAMP) is recognized by a specific pattern-recognizing receptor (PRR) on plant plasma membrane. The interaction between PAMP and PRR will trigger a variety of signalling pathways and pathogenesis-related genes. Consequently, immediate defence responses are triggered (Bigeard J et al., 2015; Mazzotta S and Kemmerling B, 2011; Povero G et al., 2011; Jones JDG and Dangl JL, 2006), frequently characterized as stomatal closure (Gudesblat GE et al., 2009), cell wall lignification (Lee MH et al., 2019), callose accumulation (Pastor V et al., 2013), hypersensitivity responses (Balint-Kurti P, 2019) and secondary metabolite production (Piasecka A et al., 2015). Chitin is a structural component of the fungal cell wall which can be recognized by a plant defence system. During plant-pathogen interaction, chitin is believed to be cleaved by plant chitinases into an oligosaccharide. The chitin oligosaccharide is recognized by plant specific PRR called “chitin elicitor receptor kinase 1 (CERK1)” which is cooperated by “chitin elicitor receptor binding protein (CEBiP)” (Yin H et al., 2016). However, some pathogens have evolved mechanisms to shield themselves from plant defence systems. One of the mechanisms is turning chitin into its deacetylated derivative, chitosan, which plant chitinases exhibited less affinity on. Some studies suggested a compound lack of a specific PRR in plants (Yin H et al., 2016). Therefore, chitosan is naturally supposed to be less efficient than chitin (Sanchez-Vallet A et al., 2015; Vander P et al., 1998). According to the results, we assumed that the defence mechanism of C. tora root cultures might not be sophisticated enough to efficiently respond to chitosan. It might have chitinases with less affinity on chitosan or consist of less compatible defence components. Meanwhile, yeast extract is a mixture of fungal cell components. It could contain many components that can trigger the defence system. Thus, yeast extract showed to be more efficient than chitosan at the same concentration. Furthermore, chitosan was suggested to negatively affect the growth and cause overstressed conditions in root tissue (Lopez-Moya F et al., 2017; Overvoorde P et al., 2010). This might be another influence that leads to the lower elicitation efficiency of chitosan in C. tora root cultures.
Both chitosan and yeast extract elicitations exhibited a concentration-dependent manner until reaching the peak level and then started to decline. This phenomenon might be explained in term of defence capacity limitation. High elicitor concentration may exceed the available number of PRRs as well as the capacity of other defence components. Moreover, overstressed conditions caused by high concentrations of pathogen elicitors might be toxic to the plant cell and decrease the overall production (Overvoorde P et al., 2010).
3.1.2 Effects of exposure time and culture age
In order to further optimise the elicitation conditions, exposure time and culture age were subsequently optimised. Our result was consistent with previous reports in Rudgea jasminoides (Oliveira MDC et al., 2007) and R. cordifolia (Ghatge S et al., 2014), suggesting that longer exposure time produced better elicitation efficiency. The highest production was observed when the root cultures were exposed to the elicitor for 48 hours. The aloe-emodin production was significantly increased from the control up to 2.93 times (0.713 ± 0.010 mg/gDW).
The culture age was another affecting factor to optimise. Without elicitation, the aloe-emodin production was drastically increased from the middle of the stationary phase to the decline phase of the growth cycle. Aloe-emodin accumulation started on day 12, gradually increased until reaching the peak level on day 20, and then plummeted after day 24. According to the production pattern, the optimisation of culture age was specifically performed during the aforementioned period – day 9, 12, 16 and 18 represented lag phase, exponential phase, late linear phase and early stationary phase of growth. Interestingly, the 100 mg/L yeast extract elicitation showed to be effective at every studied culture age. Treatment on day 12 promoted the highest elicitation benefit, with the 2.16 time-improvement in aloe-emodin production (0.1802 ± 0.012 mg/gDW) but elicitation on this day affected the root cultures health. Meanwhile, the treatment on day 16 did not show significant change when compared with the control. Interestingly, elicitation on day 18 showed negative impact on aloe-emodin production, actually lowering the aloe-emodin content to 55% of the control. (Fig. 3). The result indicated that the elicitation was the most efficient during the exponential growth phase. However, it was too early to harvest the root cultures immediately after the 48 hours contact period (day 14) because the root cultures were not fully grown. Thus, we decided to perform elicitation on day 12 and harvest the elicited root cultures at the end of the culture cycle (day 21) in the following experiments.
3.2 Simultaneous double elicitation on aloe-emodin production
In the present study, 10, 50 and 100 mg/L chitosan in combination with 100 mg/L yeast extract were used to elicit the root cultures on day 12. The aloe-emodin levels were observed at the end of the culture cycle (day 21). Without chitosan, 100 mg/L yeast extract elicitation (Y100) increased the end cycle aloe-emodin production 6.21 times (5.740 ± 0.584 mg/gDW). It was found that 10 and 50 mg/L chitosan in a combination with 100 mg/L (DC10Y100 and DC50Y100) increased the production 8.82 times (8.139 ± 0.241 mg/DW) and 9.09 times (8.389 ± 0.400 mg/gDW), respectively. In contrast, a combination of 100 mg/L chitosan and 100 mg/L yeast extract (DC100Y100) overstressed the root cultures and caused the aloe-emodin content dropped when compared with the other treatment groups (Fig. 4). With the presence of a relatively low concentration of chitosan, 100 mg/L yeast extract elicitation enhanced aloe-emodin production better than as a single elicitor. Meanwhile, the presence of the relatively high concentration of chitosan might have competitively consumed the defence resources and negatively affected the viability of the root cultures. Therefore 100 mg/L chitosan and 100 mg/L yeast extract failed to make a synergistic elicitor duo. This part of study suggested that simultaneous double elicitation of 10 mg/L chitosan and 100 mg/L yeast extract were the most effective conditions for enhancing aloe-emodin production in C. tora root cultures.
Double elicitation is a current technique to improve the efficiency of elicitation. It is based on the synergistic effect of two or more elicitors. Chitosan has been reported to synergistically work with many types of elicitors such as jasmonic acid and fungal oligosaccharide. Recently, a few studies have suggested that chitosan is involved in jasmonic acid-signalling pathway (Jia X et al, 2018; Yin H et al., 2013; Povero G et al., 2011). The combination of chitosan and methyl jasmonate exhibited a synergistic effect on glucotropaeolin production in Farsetia aegyptia (Al-Gendy AA and Lockwood GB, 2015). Interestingly, the same pathway was consistently reported to be involved with yeast extract treatment. The increase of intermediate, precursors and catalytic enzymes involved in jasmonic acid biosynthetic pathways were reportedly observed following yeast extract elicitation (Yaguchi T et al., 2017; Hasanloo T et al., 2009). Thus, chitosan and yeast extract would bi-laterally enhance the aloe-emodin production through the jasmonic acid-signalling pathway and create a new synergistic elicitor duo.
3.3 Sequential double elicitation on aloe-emodin production
Sequential double elicitation was designed based on the theory of plant priming. Priming is an adaptive strategy which enhances plant defence systems. It is a consequence of plant-pathogen interaction. Priming has been suggested to confer an underlying state for long-term immune protection in plants (Shine MB et al., 2019; Shah J and Zeier 2013; Conrath U, 2006). Consistently, plants with primed states have been reported to be more sensitised to stress and able to produce more intense defence responses (Malerba M and Cerana R, 2018; Jia X et al., 2016; Fitza KNE et al., 2013; Pastor V et al., 2013; Po-wen C et al., 2013; Prakongkha I et al., 2013; Povero G et al., 2011; Paulert R et al., 2010; M and Cerana R 2008; Katz VA et al., 1998). In this part of the study, we took advantage of the primed state to further improve the efficiency of 100 mg/L yeast extract elicitation. Chitosan showed the ability to trigger defence response in the studied root cultures. Thus, it was chosen as the priming inducer. Various concentrations of chitosan were added into the culture media on day 12 in order to prime the root cultures. After 24 hours, 100 mg/L yeast extract was added to elicit the root cultures. Aloe-emodin production was measured at the end of culture cycles. It was found that suboptimal elicitor concentrations of chitosan created an entirely different priming effect. Root cultures primed with 50 mg/L chitosan (PC50Y100) showed to be more sensitised to 100 mg/L yeast extract elicitation. However, root cultures primed with 10 mg/L chitosan (PC10Y100) were not able to elevate the aloe-emodin level when compared with 100 mg/L yeast extract elicitation alone. PC50Y100 was able to elevate the aloe-emodin production (10.401 ± 0.531 mg/gDW) up to 11.27 times higher than the control which was 1.81 times higher than using yeast extract alone. Interestingly, chitosan at 10 mg/L and 100 mg/L failed to activate the priming effect. Both root cultures primed with 10 mg/L and 100 mg/L chitosan (PC10Y100 and PC100Y100, respectively) did not improve aloe-emodin production when compared to single yeast extract elicitation (Fig. 5).
The result was consistent with many reports, suggesting that defence responses under the influence of primed states are more intense than usual. The priming effect is highly influenced by a concentration of priming inducer. As recently reported by the study in Plumbago indica root cultures, pretreatment with the effective elicitor concentration of chitosan (150 mg/L) failed to improve the elicitation effect of 100 mg/L yeast extract on plumbagin production (Jaisi A and Panichayupakaranant P, 2020). This strongly agrees with our study and emphasizes that an effective elicitor concentration of chitosan does not work as a priming inducer concentration for 100 mg/L yeast extract elicitation. Meanwhile, pretreatment with low concentrations of chitosan was reported to enhance the disease resistance against fungal infection in Pinus patula seedlings and was involved with the increased expression level of gene encoding phenylalanine ammonia lyase (PAL) (Fitza KNE et al., 2013). The improved disease resistance was suggested to be associated with the improved hydrogen peroxide level in oxidative burst (Paulert R et al., 2010). Moreover, low concentrations of systemic acquired-resistance (SAR) inducers such as benzothiadiazole, 2,6-dicholoroisonicotinic acid and salicylic acid were reported to work as priming inducers in Petroselinum crispum cell cultures (Katz VA et al., 1998; Kauss H et al., 1992). These studies support our finding that low concentration or suboptimal concentration of elicitor is required for priming and suggests that different plant species require different types and concentrations of priming inducers.
To date, the majority of priming study have been focusing on pathogen resistance outcome. Consistent reports in term of improved pathogen resistance have made priming to the practical application for crop protection (Westman SM et al., 2019; Dewen Q et al., 2017; Xiao W et al., 2017 and Iwata M, 2001). Whereas in terms of secondary metabolites, the study is completely different and needs more intensive investigation. Recently, reports on secondary metabolite production under the influence of primed state have been from parsley cell culture-based studies, using SAR-inducers as the priming inducers (Katz VA et al., 1998; Kauss H et al., 1992) and a recent study in P. indica cell cultures which used methyl-β-cyclodextrin as the priming inducer (Jaisi A and Panichayupakaranant P, 2020). Our work is probably the first to report the success of using chitosan as a priming inducer to improve secondary metabolite production. It suggests a new aspect of chitosan priming, since previous reports largely focused only on the effect on disease resistance.
The precise mechanism of priming is still vague. Nevertheless, current suggestions give priority to the pre-existing accumulation of defence molecules (signalling compounds, inactive enzymes and hormones), primary metabolism remodelling and epigenetic modifications. We assumed that these intracellular changes might be the factors that discriminate the priming effect of different concentrations of chitosan in this study. (Tugizimana F et al, 2018; Espinas N et al., 2016; Zhu QH et al., 2016; Rojas CM et et al., 2014; Conrath U, 2011; Jaskiewicz M et al., 2011). Chitosan at 10 mg/L might be insufficient to induce significant aloe-emodin production itself or to sensitise the root cultures. Thus, the production in the PC10Y100 was purely contributed by the effect of yeast extract. Likewise, 50 mg/L was insufficient for the significant elicitation in a 24 hours period, but it might be intense enough to trigger PTI and induce the root cultures to the primed state. The case of PC100Y100, which 100 mg/L chitosan worked poorly as priming inducer, could be explained in term of primary metabolism remodelling. It has been suggested that plants lose resources and energy during PTI induction due to metabolic changes, affecting functions required to maintain normal growth and living (Schwachtje J et al., 2018; Rojas CM et al., 2014). Furthermore, the balance of jasmonic acid and salicylic acid signalling pathways were also reported to be interfered, following PTI induction (Schwachtje J et al., 2018). In order to compensate the imbalance and supply the alternative resources for normal living as well as further defence responses, plants need to process primary metabolism remodelling (Schwachtje J et al., 2018; Rojas CM et al., 2014). Single elicitation of 100 mg/L chitosan significantly increased the aloe-emodin production. This implies that 100 mg/L chitosan was able to trigger PTI and lead to the depletion of cellular resources. Thus, the root cultures primed with 100 mg/L chitosan probably need some appropriate interval to process primary metabolism remodelling for the subsequent responses to 100 mg/L yeast extract. According to the result, it might be assumed that 24 hours was not enough time for the root cultures to fully process primary metabolism remodelling. Thus, priming with 100 mg/L chitosan for 24 hours prior to 100 mg/L yeast extract elicitation (PC100Y100) did not promote further improvement over the single treatment of 100 mg/L chitosan (C100). Furthermore, PAMPs have been known to promote negative effects on plant health. Chitosan, especially at high concentrations, was reported to negatively affect root growth and development (Overvoorde P et al., 2010), as well as interfere with expression patterns of phytohormones and cause overstress in root tissues (Lopez-Moya et al., 2017). Thus, exposing the relatively high concentration of chitosan following by yeast extract might cause root culture death, and possibly be involved with lower aloe-emodin production.
{kind=link}