This study investigated the intra- and inter-species variation of Amblyomma spp. collected in southern Africa. In total 7,734 adult Amblyomma ticks were collected and morphologically identified as A. eburneum, A, hebraeum, A. pomposum and A. variegatum. However, based on our analyses of multiple genes, we cannot conclude that A. pomposum and A. variegatum are distinct species.
Amblyomma eburneum is described as an eastern African species with a documented geographical distribution ranging from Somalia, Eritrea and Ethiopia in the north, and south through Kenya, Tanzania and Zimbabwe [7]. In this study we collected A. eburneum from central Mozambique in the Sofala province from African buffalo. Amblyomma hebraeum, from cattle, was collected in South Africa, Mozambique, and Zimbabwe, corresponding with its documented geographical distribution [6, 8, 10, 42]. The geographical spread of A. pomposum was described by Robinson [43], Walker and Olwage [10], and Petney et al. [42] to range from Angola to the western regions of Zambia, and northwards to the southern parts of the Democratic Republic of Congo. In the present study, Amblyomma spp. were also collected in these regions, from the western parts of Angola and latitudinally to the eastern parts of Zambia. We found A. pomposum to be restricted to the central-western parts of Angola, whereas Amblyomma specimens collected in eastern Angola were identified as A. variegatum. In this study, A. variegatum was collected in Angola, Mozambique, Zambia and Zimbabwe, corresponding with records of Petney et al. [42] and Walker and Olwage [10].
The most predominant collected species was A. hebraeum from South Africa and Mozambique, while the species with the lowest representation was A. pomposum from Angola. The low recovery of A. pomposum, and A. variegatum in Angola, could be ascribed to the collections occurring in March, which is at the end of the adult season (the months when the maximum infestations occur have been documented as November and December [42]). Amblyomma eburneum was also collected in low numbers, but this can be attributed to the difficulty of collecting ticks from wildlife species. The collections were highly dependent on the amount of wildlife that was purchased and legally hunted in the timeframe of this study. The low prevalence of Amblyomma spp. from Zimbabwe was likely a result of a vigorous campaign by the government to assist livestock farmers in treating their cattle against ticks by encouraging regular acaricde dipping of the animals, as an attempt to control major outbreaks of bovine theileriosis.
This study aimed to provide insight to the unresolved debate that has been ongoing for decades as described in the introduction. Both Robinson [26] and Dias (1950,1953) described morphological variation in the A. variegatum found in Mozambique and suggested new species names A. variegatum var. nocens and A. variegatum var. govurensis, respectively. Dias [22] also examined what was described as A. pomposum from Angola and concluded that it does not resemble the A. pomposum as described by Dönitz [25], suggesting a new species description as Amblyomma superbum. In this study, morphological variation in the collected A. variegatum ticks were observed; however, the majority resembled that of the original description by Fabricius [24]. Morphology is the most important aspect for taxonomy, highlighting the importance of variation in ornamentation. These conflicting observations of morphology in A. variegatum resemble the controversy of the Amblyomma marmoreum complex [44].
The Amblyomma marmoreum complex encompasses five African species, namely: A. marmoreum sensu stricto, A. sparsum, A. falsomarmoreum, A. nuttalli, and A. paulopunctatum [44]. As with this study, the A. marmoreum complex was placed under scrutiny due to a lack of genetic and ecological data on its members. Although meticulous descriptions of the five species in the A. marmoreum complex are available, identification remains a challenge and misidentification often occurs, even confusing these species with other Amblyomma spp. outside of the complex [23, 33, 44]. However, Cotes-Perdomo et al. [44] was able to differentiate between the species with the use of molecular techniques targeting a large part of the mitogenome. Overall, they were able to concatenate and compare 13 protein-coding genes and two ribosomal genes of several Amblyomma spp.
In the attempt to provide clarity on the phylogenetic positioning of A. pomposum and A. variegatum, molecular analyses were conducted on these four Amblyomma species with the use of 12S, 16S, coi, cytB and ITS2 molecular markers. However, the ITS2 marker was excluded from further analyses due to uninformative single nucleotide mutations. This research provides additional sequences that have been deposited in the GenBank database for all other markers utilized here, including the first for the 12S and cytB genes of A. eburneum and first for the 12S, coi and cytB genes of A. pomposum. During the course of this study, several challenges with amplification occurred for all genes. The Chelex extraction method has several drawbacks including the rapid degradation of the extracted DNA after two years of storage and the effects of long-term storage on the binding of impurities to DNA. Singh et al. [45] noted that samples extracted with the Chelex 100 resin method and stored for extended periods of time tended to contain contaminants such as proteins attached to the DNA helix, and these prevented successful PCR reactions. The importance of the impact of storage time on DNA integrity was evident through the successful amplification of cytB, which was conducted with DNA extracted less than one year previously, compared with the less successful amplification of 12S, 16S, coi and ITS2, which were conducted using DNA stored for approximately two years. Additionally, several amplified products did not generate sequences of good quality, and were thus excluded from further analysis, lowering the sample size. The 12S and 16S rRNA gene markers were particularly difficult to amplify in A. pomposum samples and those that did produce bands for these markers did not provide high-quality sequences. For future studies, it would be ideal to use recently extracted DNA when using the Chelex extraction method and DNA should be stored at -80°C, which would maintain the integrity of the DNA for a longer period; or alternative DNA extraction methods should be used that will not compromise the DNA integrity during storage.
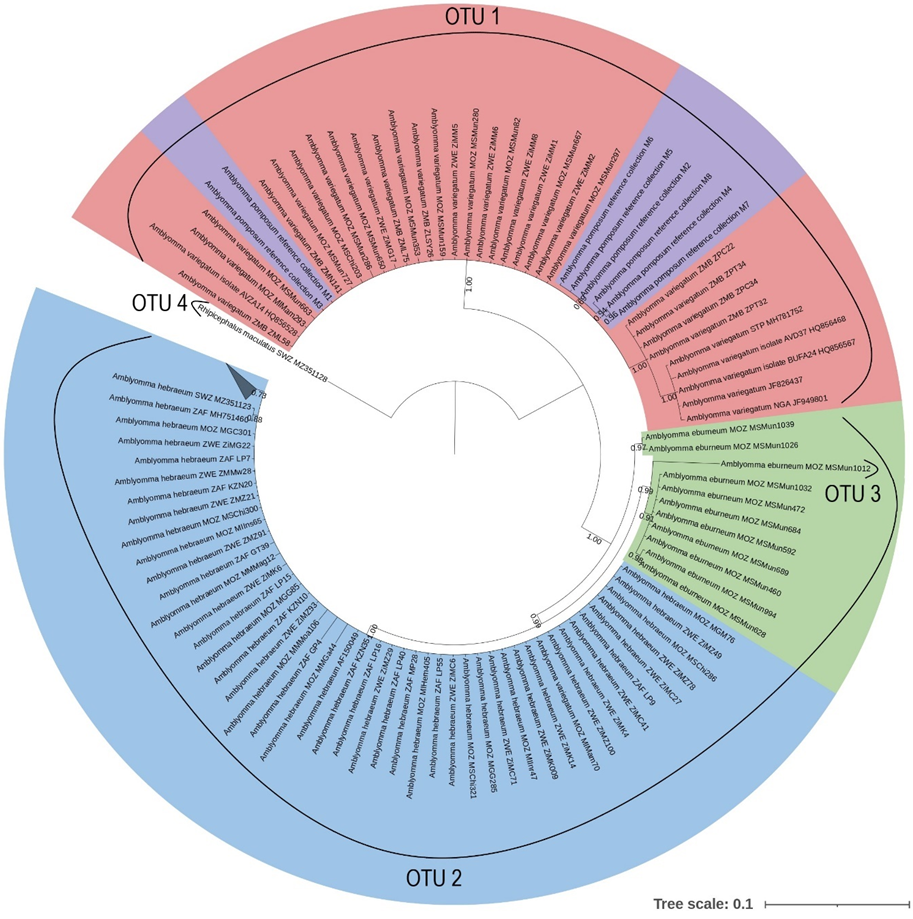
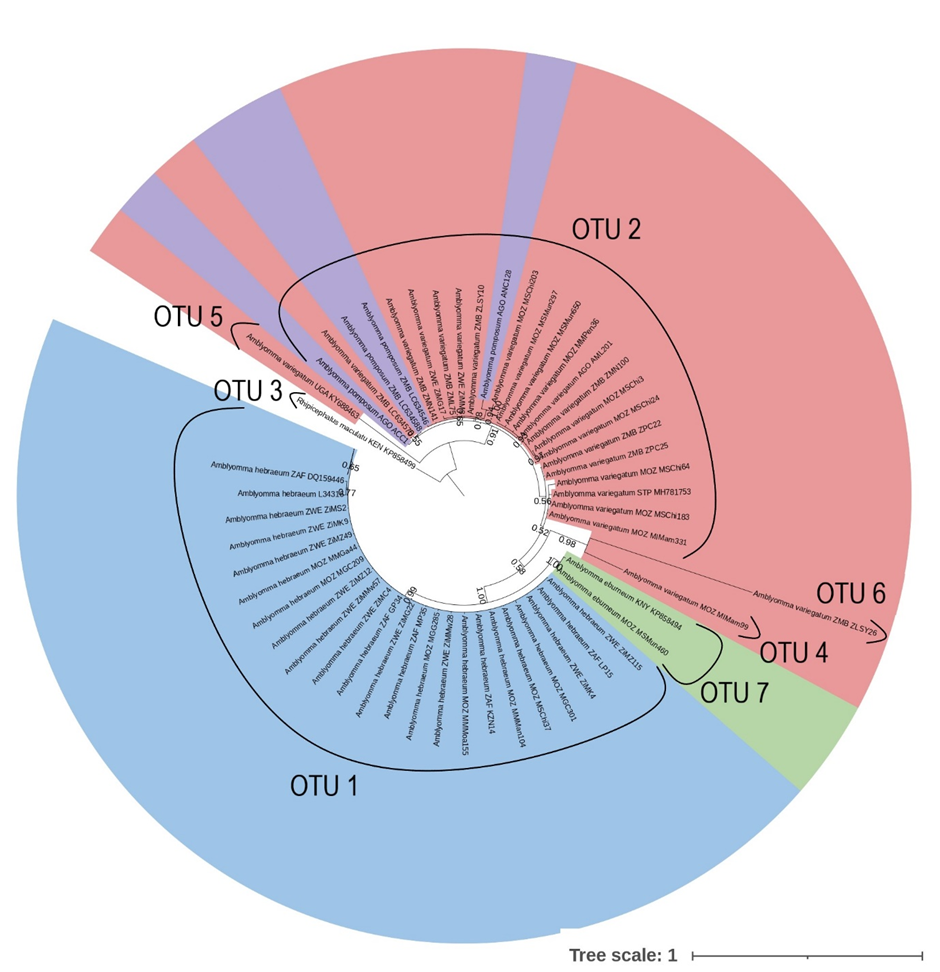
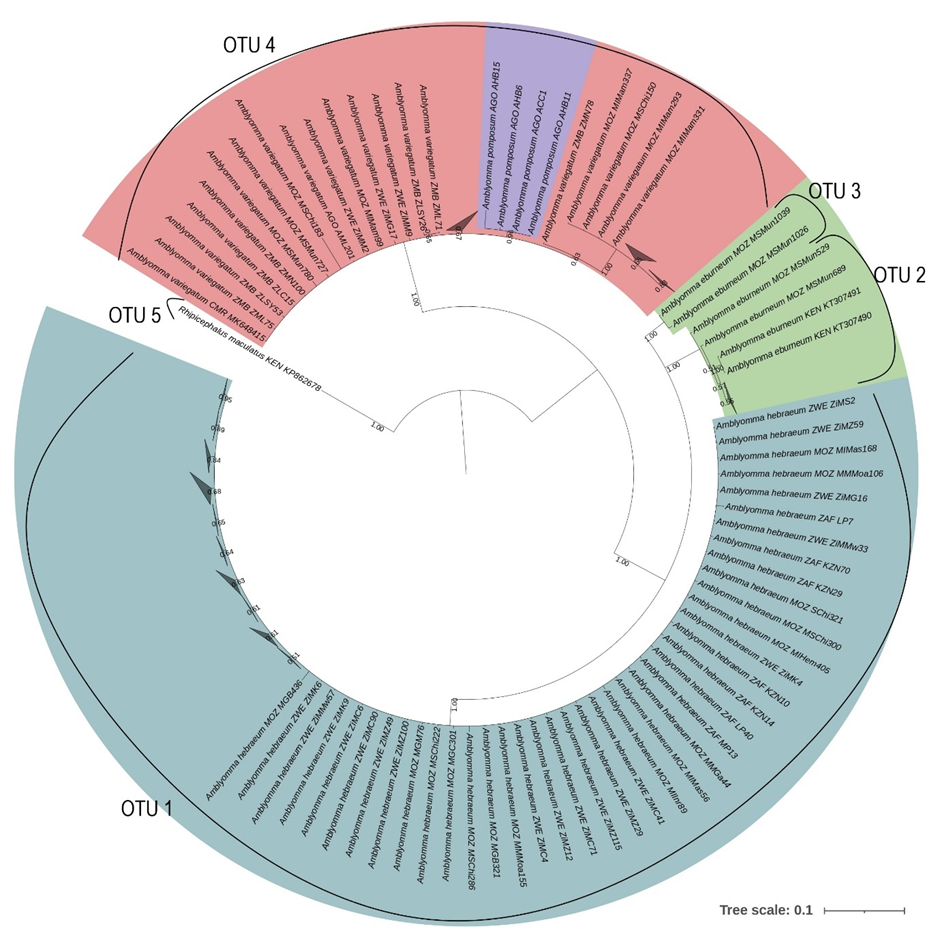
The ABGD analysis indicated that A. eburneum was divided into two OTUs in the 12S and coi analysis, while only forming one OTU in the 16S, cytB, and concatenated analyses. All the A. eburneum 12S sequences clustered with the A. hebraeum OTU, except for one which formed its own OTU. However, upon investigation, the sample that formed its own OTU was of suboptimal quality and several incongruencies was found between the forward and reverse sequences. The intraspecific variability for A. eburneum was greater for the 12S (0.018) and coi (0.021) genes, while the inter-species variability was sufficient to differentiate between species for 16S, coi, cytB and in the concatenated analyses. The inter-species variability was insufficient for the 12S analysis, as can be seen in the clustering with A. hebraeum during the ABGD analysis, though a clear separation was obtained in the phylogenetic tree. The ABGD analysis indicates that A. hebraeum only formed one OTU in all the single gene and concatenated analyses, although the intra-species variability for A. hebraeum was greater in the concatenated pairwise distance analysis. This supports the clear separation of this species from the other Amblyomma spp. of southern Africa. Strikingly, Amblyomma pomposum clustered with A. variegatum in the ABGD analyses for all the individual genes and in the concatenated analyses. The intraspecific variability in A. pomposum was greatest in the 16S and the cytB analysis. Amblyomma pomposum clustered within A. variegatum for the 12S, 16S, coi and concatenated phylogenetic analyses; however, in the cytB analysis it segregated as the ancestral lineage. The intraspecific variation within A. variegatum was greater than the interspecific variation between A. pomposum and A. variegatum for the 12S, 16S, cytB and concatenated analyses. This may indicate that there is insufficient variation between A. pomposum and A. variegatum to confidently describe these two species as distinct. The ABGD analyses for A. variegatum indicated four OTUs for the 16S and three OTUs for the cytB genes, while the 12S, coi and concatenated analyses indicated only one OTU. Two of the OTUs in the 16S analysis were the result of suboptimal sequences with several incongruencies between the forward and reverse sequences, while the other was a reference sequence from GenBank which covered a larger section of the 16S gene. As with A. pomposum, the intra-species variation for A. variegatum was greater in the 16S and cytB genes.
Phylogenetic analysis of the Amblyomma spp. indicates that all genes used in this study were adequate to differentiate between A. eburneum, A. hebraeum and the A. variegatum/A. pomposum complex. The different patterns that emerge between the phylogenetic trees could be a result of the differential mutation rates of each of the molecular markers [46]. Erster et al. [47] and Koroiva and Santana [48] evaluated the marker efficiency of 12S, 16S, coi and cytB, which were also used in the current study, and demonstrated that the mitochondrial markers coi and cytB were most suitable for intra- and inter-species analyses due to their high variability. Furthermore, Vences et al. [49] reported that cytB was the most variable of the two markers, allowing for clearer separation between closely related species. Norris et al. [50] found that 16S was the least variable marker when comparing 12S and 16S markers for Ixodes scapularis population genetics. In the current study, cytB proved to be the most efficient single marker to differentiate between southern African Amblyomma spp., while none of the markers in isolation allowed for clear discrimination between samples of the same species from different countries. However, there was strong support for intra-specific population structure in A. variegatum in the concatenated analysis.
In this context, a previous analysis of intraspecific variation of A. variegatum molecular markers concluded that genetic diversity was low in West Africa and in introduced Caribbean populations (i.e., nucleotide diversity of 0.02–0.25% for 12S), with higher variation in East Africa (i.e., 0.65% for 12S) [20]. Although the study did not apply the more variable markers as used in the present work, the 12S data alone suggests that southern populations of A. variegatum show greater genetic variation (nucleotide diversity of 1%) than elsewhere on the continent. Notably, a population genetic study of A. variegatum in Burkina Faso used microsatellites and concluded that effective population sizes were low at the village level when sampling domestic ruminants [51]. Similar approaches could also be applied to populations from southern Africa in future studies. Recently, mitochondrial markers were analysed to determine the genetic structure of another African Amblyomma spp., the elephant tick A. tholloni, on host populations in Kenya. The intra-specific variation at the coi locus was found to be low and of similar magnitude to that of A. hebraeum and A. pomposum in the current study (< 1%) [52].
Population genetic studies of Amblyomma spp. have been more extensive in the New World and offer important lessons for understanding intra-specific variation in the Afrotropical species. There are marked differences between the low intra-specific variation observed in some Neotropical species (e.g., Amblyomma triste [53] and Amblyomma aureolatum [54], where it was < 1% for mitochondrial markers) compared with others such as Amblyomma ovale [55, 56] and Amblyomma mixtum [57], where pairwise distances for concatenated mitochondrial markers can reach 3–5%. Amblyomma cajennense, which has an extensive distribution across subtropical and tropical regions of the Americas, constitutes a particularly interesting paradigm for the genus. Variation exceeding 8% at the whole mitogenome level between geographically and ecologically distinct populations of A. cajennense formed part of the evidence that was used to designate a species complex and formally describe its members as distinct species [58, 59]. The Afrotropical species await similar rigorous analyses using whole mitogenome data across the entirety of their range, in line with the recent study on the Amblyomma marmoreum complex [44].
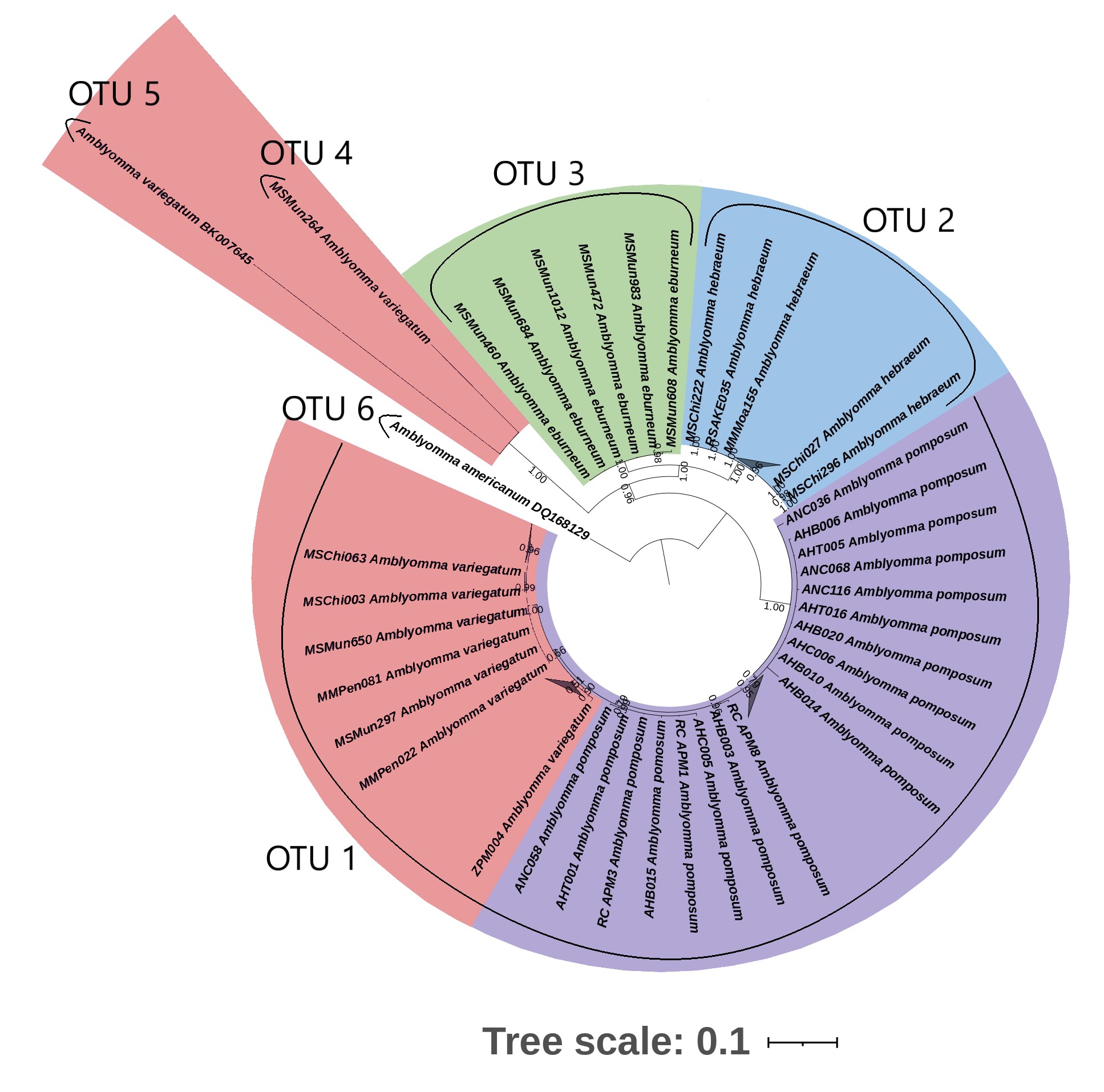
While the current study was not designed to address intraspecific population structure in depth, the concatenated phylogenetic analysis depicted clear differentiation between each of the analysed species; albeit A. pomposum branched from the main cluster of A. variegatum with a very short genetic distance, no greater than intra-specific variation within A. variegatum. An important limitation was that the concatenated tree was only constructed with two genes, coi and cytB. This was because not all loci had amplified successfully for each specimen, and also due to Koroiva and Santana [48]’s conclusion that coi and cytB markers were more suitable for intra- and inter-specific delineation. These were also the only markers that had sufficient representation amongst all the Amblyomma spp. from this study.
Thus, based on the phylogenetic analysis of the individual genes, the pairwise distance analyses and the ABGD analyses, we cannot conclude that A. pomposum and A. variegatum are distinct species. Literature on systematic work with A. pomposum is scarce and currently there are no alternative methods to compare differences or similarities between these two species, except for morphological descriptions. No previous studies on the phylogenetic relationship of these two species have been conducted and, as discussed above, there is a scarcity of A. pomposum sequences available in GenBank. Kobayashi et al. [60] were the first authors who published the only sequences currently available for the A. pomposum on the GenBank database. They collected 15 Amblyomma ticks and morphologically identified them as A. variegatum (n = 13) and A. pomposum (n = 2) with the use of Walker et al. [8]. They then conducted a phylogenetic analysis using the 16S rRNA gene. The A. pomposum that they identified clustered within the A. variegatum clade. This may suggest that the current available sequences are from misidentified A. variegatum ticks. Balinandi et al. [61] morphologically identified ticks they collected from Uganda as belonging to A. pomposum and A. variegatum; however, due to a lack of A. pomposum sequences at the time, phylogenetic analyses of the 16S gene clustered A. pomposum with A. variegatum. This discovery led the authors to believe that they had misidentified A. variegatum ticks as A. pomposum. Our 16S phylogenetic analysis depicts A. pomposum dispersed within the A. variegatum cluster. Based on our analysis of several genes, it is possible that Balinandi et al. [61] correctly identified their ticks as A. pomposum; however, since they did not upload any sequences nor depicted the morphological discrepancy, no definite conclusion can be made based on this information. On the other hand, a study performed by Barradas et al. [62] collected 116 ticks from the Huambo province in Angola. With the use of morphological identification using identification keys from Walker et al. [8] as well as molecular identification targeting the 12S and 16S rDNA genes, they identified their 11 (10%) Amblyomma ticks as A. variegatum. As with the 16S, the 12S also depicts clustering of A. pomposum within A. variegatum. In our study we collected from the same sights in Huambo and identified the Amblyomma spp. circulating in the area as A. pomposum. Our findings are also supported by Sili et al. [63], who identified all the Amblyomma spp. in the area as A. pomposum. Based on our findings we believe that Barradas et al. [62] misidentified their Amblyomma spp. as A. variegatum instead of A. pomposum.
Although the phylogenetic analyses cannot differentiate between A. variegatum and A. pomposum currently, sufficient morphological features are documented to distinguish these species from each other. The other main factor in defining a species is the ecology and biological habitats [64]. Mayr [64] stated that a species can be described as “The segregation of the total genetic variability of nature into discrete packages, so called species, which are separated from each other by reproductive barriers, prevents the production of too great a number of disharmonious, incompatible gene combinations. This is the basic biological meaning of species and this is the reason why there are discontinuities between sympatric species”. A clear parapatric boundary was observed between A. pomposum and A. variegatum; however, a hypothesis of incipient speciation has yet to be tested. We suggest, alongside whole genome phylogenetic analysis, mating and hybrid viability studies between the two species to confirm their reproductive isolation.
{kind=link}
{kind=link}
{kind=link}
{kind=link}