Organoids are self-organizing 3D cell cultures that mimic some of the cellular, structural, and functional complexity of the native organ in vitro. They provide valuable insights to understand the structure-function relationship of human organs that 2D models cannot achieve, such as brain lobe structure, renal pyramid function and heart spatial patterning. Assembloids result from the integration of multiple organoids or combination of organoids with other cell types1–3. They have the advantage of enabling the study of interaction of tissues that may not normally develop from the same progenitor, for example forebrain and hindbrain1 or vasculature and brain tissue61. A major outstanding issue in the organoid/assembloid field is the lack of innervation of peripheral tissue organoids. This is despite the fact, that almost all organs outside the brain are innervated by the peripheral nervous system, and that this neural regulation is crucial to the development, integrity, and function of organs.
Few innervated organoids have been reported to date; however, the common disadvantage of those exciting reports is that they cannot be easily adapted to other organ type organoids or are technically difficult to reproduce by researchers. Workman et al. mixed specified hPSC-derived vagal NC cells with developing gut tube organoids and created an intestinal organoid with enteric neuron innervation. This elegant work laid the conceptual groundwork for innervating peripheral tissue organoids9. However, enteric neurons are specific to the GI tract and do not innervate other organs. Thus, making this less universal as an innervation strategy for many different organoid types. Schneider et al. created a 3D bioengineered hPSC-derived CM model with autonomic innervation using hPSC-derived autonomic organoids12. CMs in this model were plated in circular form on dynamic stretch devices, which achieved advanced maturation of cardiac tissues. Autonomic neural organoids were then stuck into the ring of the engineered cardiac organoids for innervation and regulation thereof. However, these assembloids did not mimic the heart structures (ventricle/atrial patterning, heart cavity, for instance) and required specific instrumentation for their analysis. Innervated muscle models, where motor neurons connect to skeletal muscle, have been achieved using the one-pot differentiation strategy from the neuromesodermal progenitors10. This demonstrated the possibility to study complicated and anatomically distal functional coupling in organoids. However, not many organs develop their cell types and innervation from a common progenitor, such as the neuromesodermal progenitor, thus this strategy is not easily transferrable to other organoids. The assembloid technology, on the other hand, presents an ideal modular platform for individual organoid components, that is suitable for innervation of organoids2,3.
Here, we describe a strategy to address the need of a method to innervate any organ type organoid relatively easily with symNs. As an example, we created cardiac organoids that are innervated by symNs. We purposely used a simple, easy to reproduce, and relatively low-cost assembly method, that does not require bioengineering or special instrumentation, with the goal for many researchers to be able to reproduce this technique. We created the assembloids in a modular way, so that researchers can adapt them to their needs. For example, one could use the symNs to innervate other organoids, such as lung, kidney, or liver. Or one could replace the symNs and innervate the cardiac organoids with parasympathetic neurons (SSRN: https://dx.doi.org/10.2139/ssrn.4318816) or sensory neurons62,63.
Organoids or assembloids must fulfil certain criteria to be useful3,28,40. (1) 3D cultures. Several groups have generated 2D co-culture models of symN-innervated CMs in recent years. In 2016, Oh et al. demonstrated a functional coupling 2D co-culture model, using hPSC-derived symNs and mouse neonatal ventricular myocytes, and showed that the beating rate of CMs can be regulated by symNs, and can be manipulated with nicotine or optogenetic stimulation50. Later, Larsen et al. established a 2D co-culture system of neonatal ventricular myocytes and sympathetic stellate neurons from control Wistar Kyoto (WKY) and pro-hypertensive (SHR) rats. They found that hypertensive symNs were able to induce hypertensive phenotypes in healthy CMs, while healthy symNs rescued the hypertensive state in CMs51. In 2020, Winbo et al. performed the 2D co-culture model using symNs and CMs both derived from hPSC, which also displayed functional coupling regulation of CMs through symNs, inducible by nicotine49. In 2022, we described that symNs derived from iPSCs from the genetic autonomic disorder Familial Dysautonomia (FD) were hyperactive and in 2D co-cultures increased hPSC-derived CM beating29. Adding the option of a 3D organoid model to this toolset for disease modeling will increase its power for discovery of disease mechanisms and drug discovery. Our hSCAs are cultured in 3D throughout their generation and maturation stages (Fig. 1). (2) Self-organization from stem or progenitor cells. Our hSCAs are assembled by mixing day 14 symNblasts and day 7 cardiac progenitors (Fig. 1). We29,30 and others35,36 have previously shown that these progenitors will form fully differentiated and functional symNs or cardiomyocytes upon continued 2D culture. Furthermore, these symNs have been employed by us to modeled autonomic dysfunctions in Familial Dysautonomia, within the SARS-CoV-2 infection milieu, and under diabetic hyperglycemia conditions29,64,65. (3) Contain multiple cell types that mimic the native organ. We show here that the hSCAs contain multiple cardiac and symN lineage cell types. The human heart consists of multiple cardiac lineages in addition to CMs, such as endothelial cells, smooth muscle cells, and cardiac fibroblast33,34. Our hSCAs contained mature cardiac muscle fibers, T-tubules, cardiac fibroblasts, epicardial and endocardial layers, as well as atrial and ventricular CMs (Fig. 2b-d and h-i, and 5f). 4. Mimicking some structural and functional features of the native organ. hSCAs contained CM-innervating functional symNs, in which NE synthesis was detected (Fig. 3a-e). The functional coupling between symNs and CMs was observed by Ca2+ imaging and could be manipulated by nicotine and optogenetic stimulation (Fig. 3f-g).
It is becoming clearer that tissue innervation is essential for proper development, maturation and even repair of most organ tissues66–71. Accordingly, symN innervation of the heart plays an important role in development and reciprocal maturation of the tissues. In 2015, Kreipke et al. used the neurotoxin 6-hydroxydopamine (6-OHDA) to induce symN lesions in neonatal mouse hearts and found that the proliferation of CMs was increased, indicating disrupted cell cycle withdrawal due to the lack of symN innervation22. In 2021, Tampakakis et al. demonstrated similar results using a smooth muscle-specific NGF deprivation mouse model, which resulted in heart-specific symN depletion in embryonic hearts, as well as increased CM proliferation23. Interestingly, while both studies showed increased proliferation of CMs due to the absence of symN innervation, the model by Kreipke et al. showed decreased heart size after symN depletion, whereas Tampakakis et al. showed enlarged heart size. This might be due to the difference in CM density (unchanged in Kreipke et al. and increased in Tampakakis et al.) in each model and the unknown effect of symN signaling to other cell types that form the heart mass22,23. Additionally, it is believed that such regulation on developmental cardiac hypertrophy by symNs is mediated by NE and adrenergic signaling72,73. Indeed, in 2022, Kowalski et al. co-cultured mouse primary symNs with hPSC-derived CMs and showed that with symN innervation, the mature cardiac gene expression and functional cardiac activity were improved53. However, they also found that treating hPSC-derived CMs alone with isoproterenol, a β-adrenergic receptor agonist, was not sufficient to induce such maturity improvement without physical connection with symNs53, implying undiscovered mechanisms in the symN-heart axis, which may also account for the reason of different heart sizes observed in the models above (P0 symN depletion in Kreipke et al., in which embryonic innervation remained, versus symN null in the heart in Tampakakis et al.). The hSCAs described here, therefore, might be an ideal model to assess development and reciprocal maturation of symNs and CMs in cultures. Using hSCAs, we modeled early heart development. Since the exact effect of NE on CMs is not fully clarified22,23,53, we used α- and β-adrenergic receptor antagonist LAB to fully block the entire downstream target of NE (Fig. 4b). In our model, hSCA size and cell number were not altered in LAB-treated organoids compared to control (Fig. 4c). Additionally, genes for cardiac maturation decreased upon LAB treatment, indicating the importance of functional symN signaling for heart development and maturation. Future studies using hSCAs may focus on identifying the effect of other symN cofactors, such as neuropeptide Y on cardiac maturation, as well as the levels of cardiac maturation in different cardiac compartments using single cell RNA sequencing.
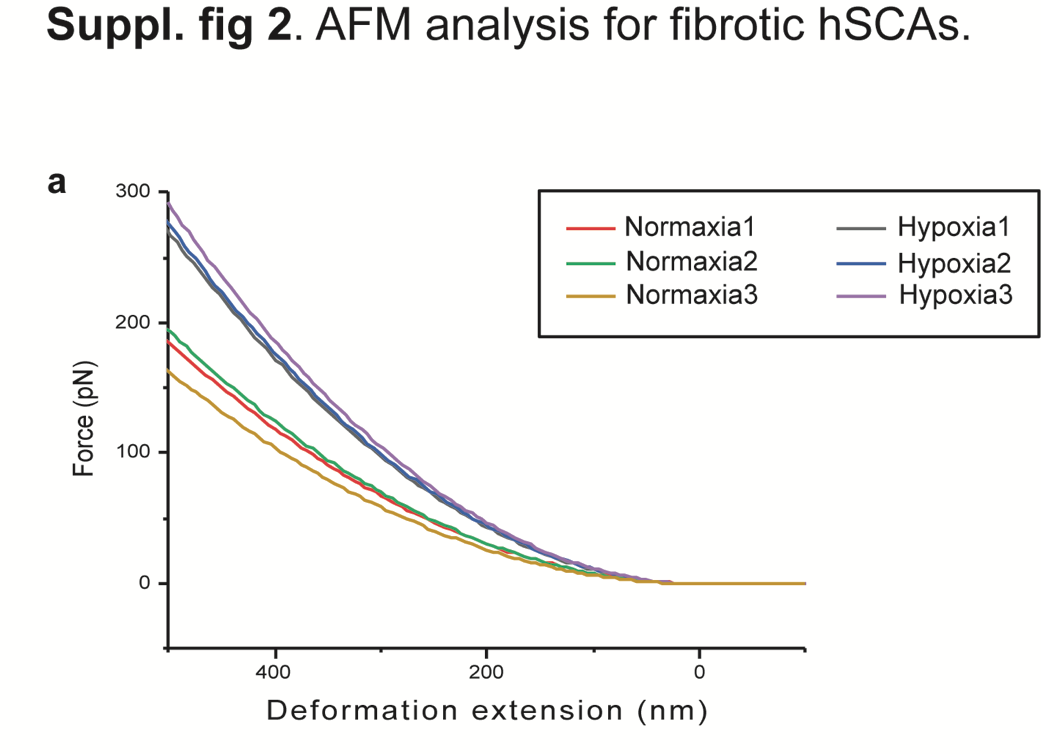
Finally, organoid technology has become an important tool for disease modeling approaches4–8,31,32,44,74. In line with such studies, we employed our hSCAs to model the hypoxia-induced cardiac infraction (Fig. 5a). We successfully recapitulated the endogenous NE crisis in hSCAs from symNs upon hypoxic stress, which caused cardiac fibrosis that was rescued by treatment with the β blocker propranolol along with the hypoxic stress (Fig. 5b-g). There are a few limitations in this model. First, in the whole organism, other NE releasing tissues, such as adrenal chromaffin cells, can also contribute to the NE crisis. Second, as a critical part during heart failure, the inflammation response was not recapitulated in hSCAs, due to the lack of immune cells in the organoids. Such challenges may be addressed in the future by incorporating more distal tissues, such as chromaffin or immune cells into a further advanced assembloid.
{kind=link}
{kind=link}