Modulation of innate immunity in Vagina Chips by mucus-containing effluents from Cervix Chips
We recently described a human Vagina Chip lined by primary human vaginal epithelium interfaced across an extracellular matrix (ECM)-coated porous membrane with underlying stromal fibroblasts that enables analysis of human host-microbiome interactions in the vaginal microenvironment,9 as well as a human Cervix Chip containing primary cervical epithelium interfaced with stromal cervical fibroblasts that produces cervical mucus with physical and chemical properties similar to those observed in vivo.10 Here, we collected mucus-containing effluents from the epithelial channel of the Cervix Chip ('cervical chip mucus' containing 4.01 ± 3.04 mg/mL of mucus glycoproteins) for 7 days and then perfused it through the epithelial channel of a Vagina Chip to simulate the natural flow of mucus in the reproductive tract in vivo (Fig. 1A). Presence of this mucus in the Vagina Chip induced statistically significant decreases in secretion of multiple relevant proinflammatory cytokines, including interleukin-1α (IL-1α), IL-1β, and macrophage inflammatory protein-1β (MIP-1β), accompanied by a concomitant increase in anti-inflammatory IL-10 protein production after 24 h of exposure compared to control chips without mucus (Fig. 1B). These results demonstrate that the mucus-containing fluids produced by human cervical epithelium in Cervix Chips in vitro can directly influence the vaginal epithelium and result in suppression of inflammatory cytokine production, even in the absence of immune cells.
Modulation of the dysbiotic vaginal microbiome by introducing cervical mucus into the Vagina Chip
We next studied the effects of cervical mucus on a dysbiotic (non-optimal) vaginal microbiome in Vagina Chips by inoculating them with a consortium containing G. vaginalis E2 and E4 combined with P. bivia BHK8 and A. vaginae (BVC1; ~105 CFU/chip) on day 14 of Vagina Chip culture in the presence or absence of mucus-containing effluents from the Cervix Chip. Interestingly, the presence of human cervical mucus inhibited the consortium's ability to colonize the epithelium and thrive on the Vagina Chip. The total number of CFU of live non-adherent bacteria collected in effluents from the epithelial channel during 72 h of infection (Fig. 2A), as well as the number of live adherent bacteria in tissue digests at the end of the 72 h culture (Fig. 2B), were significantly reduced whether Vagina Chips were pretreated with mucus effluents for 1 day before microbiome introduction, 1 day after BVC 1 addition, or continuously for the entire 3-day culture starting 1 day before the addition of bacteria.
Consistent with these data, when we quantified vaginal epithelial cells with a Clue Cell-like appearance (i.e., covered with bound bacteria)15 in digests of Vagina Chip epithelium with the dysbiotic BVC1 consortium, we observed a decrease in the number of these cells in the presence of cervical mucus (Fig. 2C,D). Not surprisingly, this reduction in bacterial cell number induced by the presence of cervical mucus was also accompanied by a concomitant increase in vaginal epithelial cell viability (retained cell number) (Fig. 2E), as well as significant downregulation of the proinflammatory cytokines, IL-8, IL-10, Rantes (CCL5), TNF-α, MIP-1β, and IL-1α after 72 h of co-culture (Fig. 3). These results demonstrate that Cervix Chip mucus can directly influence the Vaginal Chip epithelium to dampen production of inflammatory cytokines and this correlates with protection of the vaginal epithelium against injury.
Suppression of G. vaginalis growth in Vagina Chip effluents
To explore whether cervical mucus acts directly to suppress bacterial cell growth or indirectly by altering vaginal cell physiology, we next compared the growth of G. vaginalis in mucus-containing effluent samples collected from the epithelial channel of control Cervix Chips (perfused with HBSS) versus effluents from Vagina Chips that were perfused with Cervix Chip-derived mucus-containing effluent for 1 day, with the bacteria cultured directly in HBSS that was used to perfuse the apical channels of our Cervix and Vagina chips as a control. Our results demonstrate that G. vaginalis grew well in the Cervix Chip mucus in 2D culture, but growth was suppressed when cultured in effluents from the Vagina Chip perfused with similar Cervix Chip-derived mucus-containing effluent or in HBSS that lacks critical nutrients (Fig. 4A,B). Importantly, when similar studies were carried out after the addition of 50% bacterial broth to provide optimal nutrient conditions, bacterial growth was restored in the control HBSS sample, but not in the sample from the Vagina Chip exposed to Cervix Chip-derived mucus effluent (Fig. 4C, D). These findings suggest that mucus components produced by the Cervix Chip induce the cells lining the Vagina Chip to express factors that suppress G. vaginalis growth.
Cervix Chip mucus alters the vaginal secretome
To further explore the effects of cervical mucus on the vaginal epithelium, we conducted mass spectrometry analysis to compare the proteome composition of the Cervix Chip effluent before and after exposure to the Vagina Chip versus the untreated Vagina Chip effluent. Of the 1752 proteins identified (Supplementary Table 1), 103 were found to be differentially abundant as determined by fold change (|log2 fc| >= 1), padj ≤ 0.05) in Cervix Chip effluents that had passed through the Vagina Chip versus either the Cervix Chip effluent or Vagina Chip effluent alone. Significant changes in the expression of multiple proteins were observed, with 64 proteins showing increased expression (Fig. 5A) and 39 proteins showing decreased expression (Fig. 5B), with the most prominent alterations highlighted in a volcano plot (Fig. 5C). PCA analysis of the proteomics data also revealed distinct segregation among these sample groups, indicating notable changes in protein expression in effluents from Vagina Chips exposed to Cervix Chip mucus compared to those from untreated Cervix or Vagina Chips alone (Fig. 5D).
Interestingly, using STRING analysis, which incorporates both physical protein-protein interactions and functional associations from various sources (e.g., automated text mining, computational interaction predictions from co-expression, conserved genomic context, databases of interaction experiments, and curated sources of known complexes/pathways),16 we found that 3 of the 37 down-regulated proteins exhibit calcium channel inhibitor activity (PHPT1, AMBP, SLC30A1). Previous research has shown that G. vaginalis strongly induces epithelial calcium influx and contraction.17 In addition, 6 of the down-regulated proteins are ECM molecules (LGALS3BP, GPC1, AMBP, SERPING1, VASN, FBLN1, FBLN2), which may play a role in G. vaginalis adhesion and biofilm formation.18 Finally, 3 down-regulated proteins are members of the Lipocalin family (AMBP, APOD, RBP4), which is known for its role in regulating inflammation and antioxidant responses.19
The STRING analysis additionally showed that 17 of the up-regulated proteins are RNA binding proteins (RBPs). Previous studies have highlighted the vital role of RBPs in bacterial replication by binding to and regulating their RNAs.20 These proteins also play a crucial role in immune system response to viral infections by regulating viral RNA stability and translation.21 Considering that BV increases susceptibility to sexually transmitted infections, including viral infections, these findings further support the potential involvement of RBPs in the immune response within the reproductive tract. Twenty-five proteins related to the male reproductive system were also found to be upregulated. This finding is significant because past studies have established a notable link between BV and infertility.6 For instance, one of the proteins identified, CSTF2T, has the potential to contribute to sperm adhesion to the zona pellucida22 while the also identified TMED10 protein may be involved in in sperm capacitation and the acrosome reaction.23
Importantly, exposure of the Vagina Chip to cervical mucus also resulted in enhanced production of potential antimicrobial proteins PLAU and WASF2. PLAU is a serine protease with immunomodulatory functions24 and WASF2 is a member of the Wiskott-Aldrich syndrome protein family that regulates autophagy and inflammasome activity.25 One of the prominent down-regulated proteins, GNS, is an N-acetylglucosamine-6-sulfatase. This is interesting because BV is often associated with the breakdown of mucins, which are necessary for these dysbiotic bacteria to colonize the vagina.26 Thus, downregulation of GNS could contribute to the inhibition of dysbiotic bacterial growth we observed by increasing glycoprotein sulfation and thereby prevent mucin degradation.
Potential role of exosomes as mediators of the effects of cervical mucus on the Vagina Chip
Exosomes, which are small extracellular vesicles containing nucleic acids, lipids, and proteins, play a significant role in intercellular communication in the female reproductive tract by modulating the immune system and promoting tissue repair.27 This is accomplished by presenting antigenic peptides, regulating gene expression through exosomal miRNA, and inducing differential signaling through exosomal surface ligands. Importantly, when we carried out STRING analysis of the proteins differentially expressed in Vagina Chip effluents exposed to Cervix Chip mucus, we found that a significant proportion of the differentially expressed proteins were associated with exosomes. Specifically, 23 out of 37 down-regulated proteins and 17 out of 64 upregulated proteins were found to be linked to extracellular exosomes (Supplementary Table 2). Notable among the upregulated proteins were DDR128 and COMP,29 which regulate cellular adhesion to the ECM and its remodeling, subsequently influencing bacterial adhesion.30 Conversely, among the down-regulated proteins, 5 ECM proteins (AMBP, FBLN1, GPC1, LGALS3BP, and SERPING1) were identified, which may also influence bacterial adhesion. Interestingly, SERPING1 functions as a regulator of the complement system31, and three of the down-regulated proteins (AMBP, SERPING1, and SPINT1) belong to the Kunitz family of serine protease inhibitors that are involved in coordinating inflammation.32
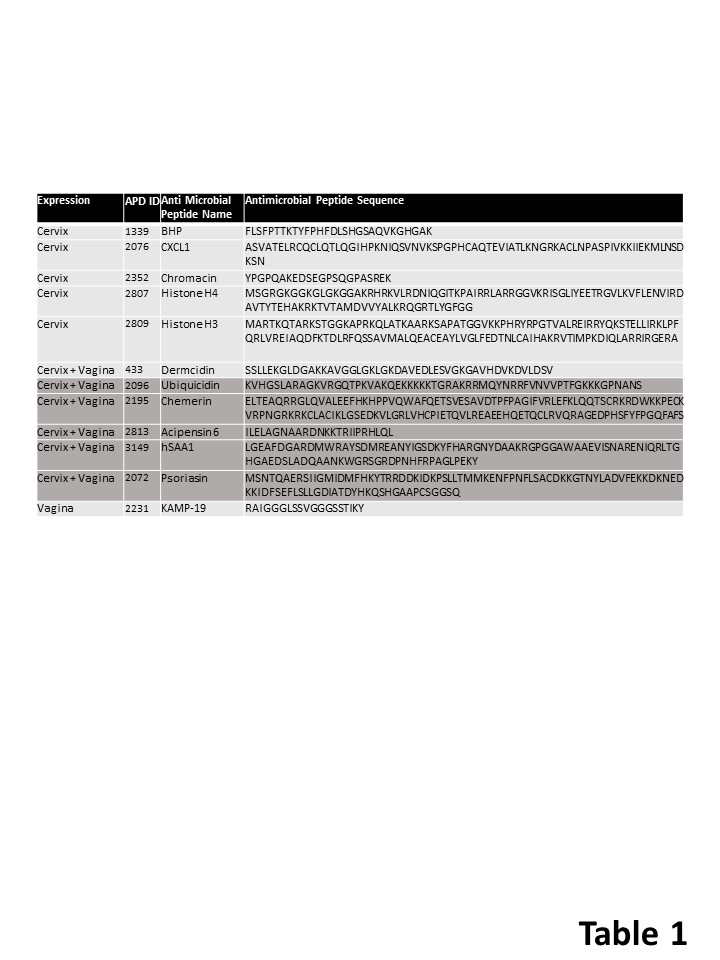
Cervicovaginal antimicrobial peptides
Additionally, we identified 12 antimicrobial peptides in the Cervix Chip and Vagina Chip effluents (Table 1). Of these, 6 (Dermcidin, Ubiquicidin, Chemerin, Acipensin 6, hSAA1, and Psoriasin) were present in both Vagina and Cervix Chip effluents, 1 was solely produced by the Vagina Chip (KAMP-19), and 5 were exclusively produced by the Cervix Chip. No antimicrobial peptides were specifically induced in Vagina Chips exposed to Cervix Chip effluents. The Cervix Chip-derived antimicrobial peptides include Histone H4, Histone H3, CXCL1, BHP, and Chromacin. Histones and their fragments have a variety of antimicrobial actions and functions, including bacterial cell membrane permeabilization, penetration into the membrane followed by binding to bacterial DNA and/or RNA, binding to bacterial lipopolysaccharide (LPS) and neutralizing its toxicity, and entrapping pathogens as a component of neutrophil extracellular traps.33, 34 It is noteworthy that P. bivia, which is included in our bacterial consortium, has been shown to produce high concentrations of LPS.35 CXCL1 also inhibits the growth of E. coli and S. aureus in vitro36 and BHP impedes growth of M. luteus, S. epidermidis, and several fungi (e.g., C. albicans, S. cerevisiae, and A. nidulans)37, while Chromacin suppresses the growth of Bacillus megaterium and Micrococcus luteus.38
Comment
Principal Findings: These data show that mucus-containing effluents from human Cervix Chips suppress the growth of dysbiotic microbiota, associated inflammation, and epithelial cell injury in human Vagina Chips. By analyzing the differentially abundant proteins in the secretome of Vagina Chips following treatment with Cervix Chip effluents, we identified multiple proteins that may contribute to this protective response and that potentially could be used as clinical biomarkers for monitoring female reproductive tract health.
Results in the Context of What is Known: Maintaining homeostasis is crucial for the health of epithelial barriers, which can be disrupted during infection or injury. Inflammation plays a vital role in supporting the body's defense against pathogens, promoting tissue healing, and restoring homeostasis.39 However, chronically high levels of proinflammatory cytokines that undermine normal protective immune signals have been linked to an imbalanced microbiome and compromised epithelial cell stability.40 This study presents evidence that communication between cervical and vaginal tissues in the lower reproductive tract via transfer of cervical mucus-containing secretions helps to suppress vaginal inflammation in the presence of a dysbiotic microbiome. There is a growing body of clinical evidence suggesting that medical cervical procedures may disrupt this crucial communication between cervical and vaginal epithelium, and lead to changes in composition of the vaginal microbiome.41, 42 Our results support this observation and suggest that it is a direct effect of reducing cervical mucus transfer to the vagina, which could only be studied directly using this type of engineered in vitro model.
The recurrence of abnormal vaginal flora after treatment of BV (e.g., with metronidazole) is commonly detected in most women,8 however, the underlying factors contributing to these recurrences remain elusive. Our findings suggest that alterations in cervical mucus levels may influence the susceptibility of the vaginal epithelium to BV infection. Therefore, an imbalance in the cervicovaginal mucus may be a possible contributing factor to the high rate of BV recurrence. In this context, it is important to note that we identified five cervical antimicrobial peptides that appear to play a role in the antimicrobial effects we observed on-chip. These findings suggest that interactions between antimicrobial peptides and the host vaginal epithelium can enhance innate immune protection against dysbiotic flora.
Immune effectors and specialized stromal cells at epithelial surfaces produce cytokines and antimicrobial defenses to orchestrate tissue repair and minimize opportunistic infections. Exosomes can act as mediators for this form of inter-tissue communication. We identified 40 exosomal proteins produced by vaginal epithelium that were modulated by exposure to cervical mucus produced in the human Cervix Chip. Human cervicovaginal exosomes have been previously shown to be part of the female innate defense system and to protect against HIV-1 infection43 as well as bacterial toxins.44 Exosomes are also currently being explored as potential therapeutic agents and drug delivery vehicles. Thus, the ability to study the role of exosomes in host-microbiome interactions in the female reproductive tract in vitro using the human Organ Chip models described here may facilitate the development of novel treatments for vaginal dysbiosis as well as other diseases of the female reproductive tract.
Clinical Implications: Our study has important clinical implications as it has the potential to identify new targets for diagnosis and treatment of vaginal diseases. Identifying patients with a high likelihood of recurrent vaginal dysbiosis can help to customize their treatment plan and prevent complications. In this study, we identified multiple proteins and antimicrobial peptides that may contribute to the protective response against dysbiotic microbiota and associated inflammation and injury to the vaginal epithelium. These proteins and peptides could potentially be used as clinical biomarkers for monitoring the health of the female reproductive tract in the future. Several proteins we identified (e.g., TPM3, PLAU, ALDH3A2, GAS6, DTYMK, SERPING, STAT6, CMPK1) are known to be targeted by existing approved drugs (Progesterone, Urokinase, Disulfiram, Warfarin, Zidovudine, Rhucin, Indomethacin, and Gemcitabine, respectively). Thus, if these molecules actively contribute to the BV disease phenotype, one or more of these therapeutics could be added to current clinical regimens.
Research Implications: Our results show the value of human Organ Chip technology for studying vaginal health and diseases of the female reproductive tract. However, further research is needed to evaluate the effects of these compounds as well as modulators of the other putative targets we identified for maintaining vaginal homeostasis and a healthy microbiome.
Strengths and Limitations: While the human Vagina and Cervix Chips used in this study replicate many physiological and pathophysiological features of the female reproductive tract, we did not incorporate immune cells. As these cells play a crucial role in mounting antibacterial immune responses, the model would be strengthened by incorporating them in the future. Additionally, it should be noted that the Organ Chips we used were created with epithelial cells from a single human donor and thus, these studies should be extended to include chips lined by cells from multiple donors from different ethnic groups as well.
{kind=link}