The outcome of this study revealed that by triggering various mechanisms, the growth-promoting endophytic fungus effectively imparted water deficit tolerance in maize. The scenario in which global climate change is emerging and significant temperature rises lead to drought stress, which reduces agricultural productivity, served as a rationale for the current study. Climate change also had a severe detrimental influence on the growth and production of several significant crops, such as Zea mays. In such a range of adverse challenges, innovative techniques and the application of sustainable eco-friendly practices can assist in breaking the feedforward loop by increasing resource use efficiency and promoting healthy crop growth while monitoring hazardous environmental conditions and enhancing soil health (Drost et al. 2020). To alleviate stress, researchers used an array of strategies, including endophytic microbes (Aziz et al. 2021; Khan et al. 2022). The implementation of such an approach is of relevance in conceptualizing plant response to water deficit stress and in gaining knowledge for the success of crop production. By providing agronomic and acclimating benefits to the host plants, microbes, especially endophytic fungi, play an important role in reducing water deficit stress.
Roughly 70 species that make up the genus Fusarium are very diverse in terms of their genetic makeup, ecology, and hence secondary metabolism. It has been isolated from a broad range of plant species and habitats. The naturally occurring endophyte Fusarium equiseti can colonize an array of hosts in different habitats and promote plant growth (Leslie and Summerell 2006; Saldajeno and Hyakumachi 2011). It could successfully colonize a plant in a range of ecosystems, which suggests that it can maintain stable populations within the host by competitively substituting pathogenic and other rhizosphere-residing organisms. Thus, it has the potential to be employed as a sort of biocontrol to enhance plant development and health (Sisic et al. 2017).
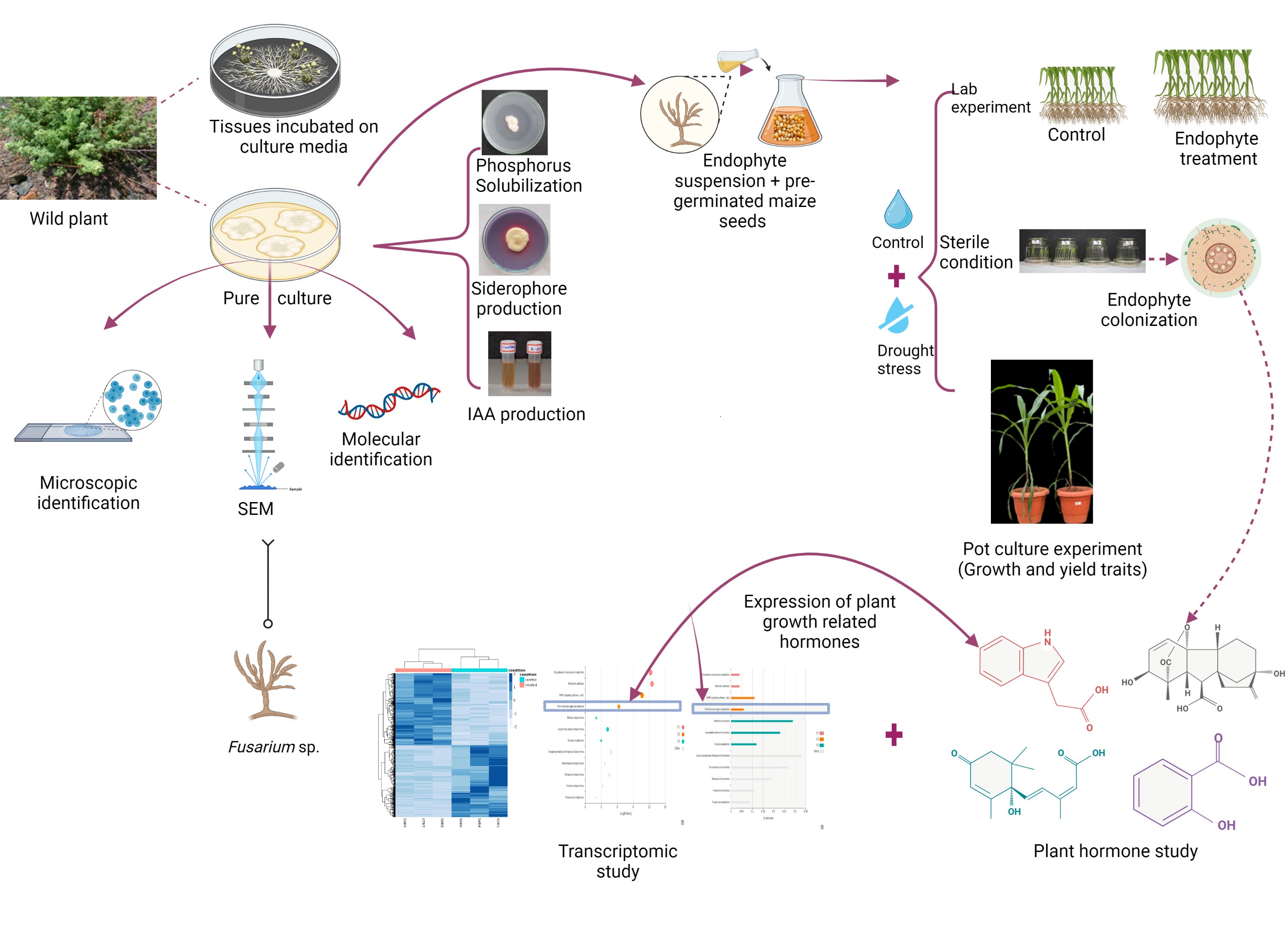
Additionally, it has long been believed that endophytes that exert phosphorous solubilization, siderophore, and IAA production are the best candidates for promoting plant growth, which boosts plant vigour and growth as it controls their developmental process (Sehar et al. 2022). The endophytic strain K23-FE was tested for its capacity to solubilize phosphate, and we observed that it could solubilize within 5 days of incubation. This outcome is comparable to a prior work by Chen and Liu (2019), where the P-solubilizing rate was maximum around day 5 in the culture condition. There are shreds of evidence on the use of phosphate solubilizing endophytes to enhance their availability to plants in soil containing insoluble phosphates (Goranada et al. 2018). Ortho-phosphate, which is accessible for plant uptake and use is formed when phosphate complexes are solubilized by fungal endophytes and released into the soil. Therefore, one crucial mechanism that might promote the development of maize plants is solubilizing phosphate by fungal endophytes. In contrast, specific microbes produce siderophores, a class of iron-chelating compounds that adhere to the ferric ion in the root zone. The ability of iron absorption by the root of some plants is boosted by the production of the siderophore-iron complex increasing plant growth (Márquez et al. 2020). In the present study, the endophytic strain K23-FE produces a siderophore, supporting its potential application as a maize plant growth stimulant. IAA synthesis by plant-associated microbes has been extensively documented (Bunsangiam et al. 2019), although fungi are less well recognized for it. Our endophytic fungus isolated from the plants adapted to extreme habitats was able to secrete IAA enriched with L-tryptophan. Similar results were noticed by a strain of Fusarium sp. which synthesized IAA (Tsavkelova et al. 2012).
In the current work, we noticed that inoculation with fungal endophyte enhanced growth in plants under water deficit stress, resulting in higher plant biomass and yield than non-inoculated ones (Table 1). Furthermore, all plants recovered irrespective of treatment when 100% FC was given the following day after the stress period. According to Golparyan et al. (2018), the most often documented plant response to endophyte inoculation in numerous plant species is an increase in seedling length. In addition to modulating host gene expression and other secondary metabolic pathways, endophytes also produce several metabolites, such as secondary compounds and plant growth hormones (Ramos et al. 2018; He et al. 2021). Similar findings were made in prior research, where under PEG-8000 induced osmotic stress, IR-64 paddy seedlings inoculated with endophytic Aspergillus sp. increased root length compared to untreated seedlings (Sangamesh et al. 2018).
In total, endophyte inoculated plants performed better in greenhouse conditions 30 days after sowing than non-inoculated plants. The growth promotion may be attributed to the increase in photosynthetic efficiency upon endophyte treatment (Figs. 3 and 4). The secretion of phytohormones and secondary metabolites by endophytic fungi, however, has been shown to be responsible for the promotion of plant growth (Ismail et al. 2021). Endophytes may also have contributed to this by helping the host plant to absorb nutrients from the soil, such as nitrogen and phosphorus (García-Latorre et al. 2021), and endophytes might have genes that assist in producing some signaling molecules for growth promotion of plants (Ali et al. 2014).
Under 100% field capacity, endophytic strain K23-FE showed a significant increase (Figs. 3 and 4) among the treatments, in photosynthetic assimilation rate, stomatal conductance, and transpiration rate compared to non-inoculated plants at 30 DAS. Plant height, root volume and dry biomass obtained also showed a similar trend with respect to photosynthetic parameters in plants inoculated with endophyte. The increase in photosynthetic efficiency of plants obtained in this study agrees with the increased photosynthetic rate, chlorophyll content, and stomatal conductance of maize plants treated with endophyte Trichoderma harzianum (Doni et al. 2019). Similar results with endophyte inoculation have been documented in earlier studies (Xu et al. 2021; Siddique et al. 2022).
The results of this study indicate that a decrease in chlorophyll concentration correlates with water deficit stress. It might be an indicator of oxidative stress caused by chlorophyll breakdown and pigment photooxidation. Because chlorophyll is crucial for photosynthesis, any decrease in chlorophyll concentration would influence photosynthetic ability. However, endophyte-treated plants retained more chlorophyll than non-inoculated plants, indicating that endophytes can increase chlorophyll stability under water deficit conditions. In endophyte-infected sunchoke plants, the rate of chlorophyll was increased (Suebrasri et al. 2020).
A substantial amount of water in plants is crucial for maintaining the osmotic levels in a plant cell, nutrient uptake, and for survival under drought stress conditions (Soltys-Kalina et al. 2016). Increased levels of RWC in endophyte treated plants at 50% FC (Fig. 5) shows that endophyte helps in maintaining higher plant water status required for physiological activities promoting plant growth and development (Touchette et al. 2014; Naveed et al. 2014; Odokonyero et al. 2016).
Small metabolites that scavenge ROS, such as proline, maintain redox equilibrium in cells undergoing oxidative stress (Meena et al. 2019). Endophyte treatment enhances proline concentration that usually favours ROS scavenging in the plants (Shavalikohshori et al. 2020). In our present study, proline was seen in an increasing trend in endophytes inoculated and non-inoculated plants under 100 and 50% FC (Fig. 5). These findings are consistent with recent reports on the ability of fungal endophytes to increase proline content under salt and water deficit stress (Moghaddam et al. 2021). An increase in ROS levels leads to elevated lipid peroxidation levels in cells resulting in membrane damage (Juan et al. 2021). MDA levels increased significantly under 50% FC compared to 100% FC in non-inoculated plants. But, in endophyte inoculated plants, there was a significant reduction in MDA levels under 50% FC compared to non-inoculated plants (Fig. 5). Endophytes elevate plant resilience to water deficit stress by maintaining proline content in host tissue and reducing MDA levels (Sadeghi et al. 2020; Zhou et al. 2021).
In this study, it was observed that the catalase and polyphenol oxidase activity was significantly high in endophyte inoculated plants under 50% FC compared to non-inoculated plants (Fig. 5). The increased activity of CAT and PPO shows the effective scavenging of H2O2 generated in endophyte treated plants under water deficit stress. This shows that endophytes in symbiotic associations are likely to upregulate the activities of CAT and PPO to scavenge ROS, aiding plants in surviving water deficit stress. Similar results have been obtained in various crop plants under water deficit stress in endophytic fungal treatments (Lata et al. 2018; Zhou et al. 2021; Siddique et al. 2022).
Thus, an increase in seed yield upon endophyte treatment could be due to an increase in net photosynthetic rate (Fig. 4) and the rate of photoassimilate remobilization from source to sink. Trichoderma sp., a fungus that inhabits the roots of stressed plants, increases yields by triggering metabolic mechanisms that modify the harmful reactive oxygen species formed under stress into less toxic molecules (Harman et al. 2021). In another study, tomato yield increased upon cross-infecting with endophytic fungi Ampelomyces sp. and Penicillium sp. (Morsy et al. 2020).
Phytohormones are well-known for their role in regulating plants’ physiological and metabolic behavior via adaptive responses under stressful situations. The current work demonstrated the ability of endophytic strain K23-FE to synthesize IAA, GA, and SA, indicating its potential function in the activation of growth responses in plant systems with decreased ABA levels. Endophytes play a pivotal role in the growth and development of host plants, in a specific manner, by secreting various phytohormones or stimulating the synthesis of native plant hormones and signaling molecules (Pinski et al. 2019; Javed et al. 2022), and thus aiding in the mitigation of various environmental factors (Shade et al. 2017).
The activation of antioxidant enzyme genes in M. oleifera plants after endophyte inoculation implies an impact on drought stress tolerance as a major regulatory function (Rehman et al. 2022). The current study found that maize plants colonized by the endophyte revealed an increase in genes linked with plant development and resilience to abiotic stress under control and osmotic stress conditions. Among the genes that have been upregulated include beta-amylase, aquaporins, LEA proteins, and transcription factors linked to plant growth hormones. Dehydrins, which play a role in stress response, are also among the elevated genes.
Here, the maize plants enriched with endophyte have been shown to differentially regulate genes involved in secondary metabolites, plant growth hormones, MAPK signaling genes, antioxidant biosynthesis, photosynthesis and carbohydrate metabolism genes. Though secondary metabolites are not directly linked to plant growth, they do help the plants well in advance to adapt to different levels of biotic and abiotic stress (Bajaj et al. 2018; Khare et al. 2020). Similarly, the medicinal plants that were enriched with endophyte were reported to have more amount of secondary metabolite accumulation when compared to untreated plants (Ogbe et al. 2020).
As shown in (Supplementary Figure S4), endophyte enriched plants have shown significantly higher seedling growth when compared to the control. Interestingly, from the transcriptome data (Fig. 9) endophyte enriched plants were shown to upregulate genes that govern the biosynthesis of growth-promoting hormones such as auxin, gibberellin, and cytokinin. In the past few years’ endophytes have been gaining much importance due to their ability to modulate growth hormones. For instance, transcriptome analysis of Achnatherum inebrians plants enriched with Epichloe endophyte has accumulated more amount of growth hormones such as GA and cytokinin (Zhao et al. 2021). Mitogen-activated protein kinases are the signaling molecules that transduce and activate phosphorylation of downstream-regulated genes such as hormone biosynthesis and signaling genes (Lin et al. 2021). Fusarium equiseti has also been shown to upregulate several antioxidant genes such as glutathione, ascorbate, etc. Endophytes enriched plants were proven to improve antioxidants, minimize reactive oxygen molecules produced, and thus lowering oxidative stress damage (Nagabhyru et al. 2022; Byregowda et al. 2022; Kaur and Saxena 2023).
Endophytic strain K23-FE has not only promoted seedling growth under control conditions but also has imparted osmotic stress tolerance to maize seedlings. RNA seq analysis of endophyte inoculated and untreated maize seedlings under osmotic stress proved the expression of genes regulating sucrose and starch catabolism. Sucrose and starch serve as precursor molecules of glucose to synthesize osmolytes such as trehalose (disaccharide) and mannitol (sugar alcohol). During stress conditions, endophyte enriched plants tend to accumulate more amount of osmolyte and thereby reducing the water potential of the plants compared to its surrounding. Endophyte enriched N. benthamiana plants exhibited enhanced relative water content, soluble sugars, and starch content when compared to endophyte-free plants (Dastogeer et al. 2018). Maize seedlings inoculated with Pseudomonas putida strain FBKV2 showed upregulation of genes associated with choline synthesis, heat shock proteins, β-alanine synthesis, and late embryogenesis abundant (LEA) proteins where all of which play a crucial role in drought tolerance (SkZ et al. 2018).
Interestingly, we found a correlation between the altered gene expression (RNA Seq study) upon endophyte treatment with the performance of maize (lab and pot experiments), especially in the early stages of the life cycle. The increased plant growth, photosynthetic assimilation rate, relative water content, proline, antioxidants (catalase & polyphenol oxidase), and reduced malondialdehyde content analogized with RNA Seq data corresponding to upregulation of plant growth hormone-related transcription factors along with beta-amylase, aquaporins, LEA proteins, dehydrins that provide stress-responsive function and antioxidant enzymatic genes.
In summary, the positive implications of endophytic fungi in maize plants have been proven by comparing plant growth, chlorophyll content, photosynthetic parameters, antioxidant enzyme activities, proline content, hormones, and stress-responsive genes in endophyte inoculated and untreated plants under water deficit stress. In short, endophyte inoculated plants showed remarkable tolerance to water deficit stress, which might be used to improve maize cultivation in dryland areas. We infer that such endophytes have evolved strategies and mechanisms that allow them to endure and benefit their host plants to thrive under difficult climate conditions. Here, the endophytic strain K23-FE isolated from plants acclimated to harsh environments was able to colonize maize seedlings while also promoting plant growth and impacting their ability to withstand water deficit stress. Therefore, endophytes could serve as an alternative approach for imparting drought stress tolerance at the field level.
{kind=link}