A. muciniphiladelayed the progression of pre-DM
Before intervention, no significant differences in FPG levels were observed between the prediabetic groups and the control group (p > 0.05). The 2h-PG levels were markedly higher in all prediabetic groups compared with the control group (p < 0.01), and there was no discernable variance between the prediabetic groups (p > 0.05) (Table 2). Following continuous intake of an HFD, FPG and 2h-PG levels in the rats with pre-DM were increased. Specifically, FPG levels significantly surpassed those in the control group (p < 0.05), and the 2h-PG levels exceeded 11.1 mmol/L, which was significantly higher than that in the other prediabetic groups (p < 0.01). However, following treatment with A. muciniphila or with dietary intervention, no significant differences in FPG levels were observed between the prediabetic groups and the control group (p > 0.05), and the 2h-PG levels observed in the prediabetic groups were within 7.8–11.1 mmol/L (Fig. 1B-D). Both treatment methods notably reduced the 120 min AUCG levels in the rats with pre-DM (p < 0.05) (Fig. 1E). These results suggest that A. muciniphila supplementation and dietary intervention are both effective methods for delaying the progression of pre-DM. Furthermore, A. muciniphila can also improve glucose metabolism in the rats with pre-DM under the condition of continuous HFD.
A. muciniphilaimproved first-phase insulin secretion and IR in the rats with pre-DM
While none of the prediabetic groups displayed a significant difference in 120 min AUCINS values (p > 0.05), the ΔI30/ΔG30 levels in the prediabetic groups treated with A. muciniphila and a chow diet were significantly higher than those in the HFD group (p < 0.05) (Fig. 1F, H, I). Both A. muciniphila treatment and dietary intervention significantly reduced the FINS and HOMA-IR levels in the rats with pre-DM (p < 0.05 or p < 0.01). With respect to the HFD group, the CD + AKK group and the HFD + AKK group exhibited more significant reductions in FINS and HOMA-IR levels than the CD group (p < 0.01) (Fig. 1G, J, K). These findings suggest that both A. muciniphila treatment and dietary intervention can increase first-phase insulin secretion, reduce hyperinsulinemia, and alleviate IR in the rats with pre-DM. Moreover, A. muciniphila supplementation can improve hyperinsulinemia and IR independently of diet.
A. muciniphilacan promote insulin secretion and inhibit apoptosis and dedifferentiation in the β cells in rats with pre-DM
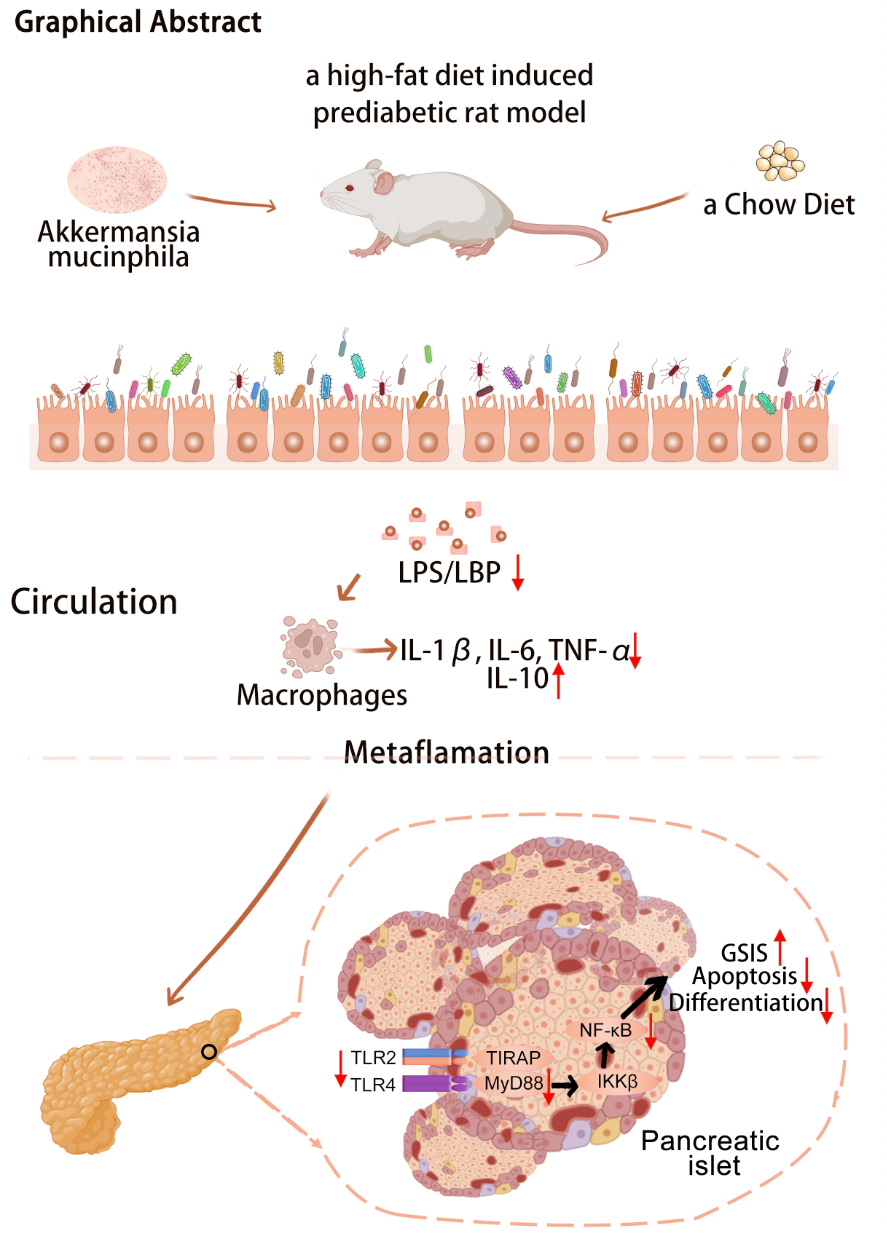
To assess the effects of A. muciniphila on insulin secretion, apoptosis, and differentiation in the β cells in rats with pre-DM, pancreatic islets and tissues were isolated from the rats in each group after 5 weeks A. muciniphila supplementation and dietary intervention. GSIS assays and TUNEL staining were performed, and the gene and protein expression of specific transcription factors related to β-cell apoptosis and differentiation was quantified. Following A. muciniphila supplementation or dietary intervention, the GSIS index in the rats with pre-DM surpassed that in the HFD group, with the more substantial increases in GSIS indices observed in the CD + AKK and HFD + AKK groups (p < 0.05) (Fig. 2A). These findings suggest that while both A. muciniphila treatment and dietary intervention can improve GSIS in the β cells in rats with pre-DM, A. muciniphila treatment can restore GSIS independently of diet.
Both A. muciniphila treatment and dietary intervention significantly reduced the percentage of TUNEL+ β cells in the pancreatic tissues of rats with pre-DM, and the more notable reductions were observed in the CD + AKK and HFD + AKK groups (p < 0.01) (Fig. 2B, C). Furthermore, A. muciniphila supplementation and dietary intervention both downregulated the mRNA expression of the proapoptotic factor Caspase3 in the islets of rats with pre-DM (p < 0.01) (Fig. 2D). Upregulated mRNA expression of the antiapoptotic factor Bcl-2 in the islets of rats with pre-DM was observed after both A. muciniphila supplementation and dietary intervention, and significantly elevated expression levels were observed in the CD + AKK group compared with the HFD group (p < 0.01) (Fig. 2E). These findings suggest that A. muciniphila treatment and dietary intervention can both inhibit β-cell apoptosis in the rats with pre-DM, while A. muciniphila can exert antiapoptotic effects on β cells independently of diet. Moreover, treatment combined with A. muciniphila and a chow diet had a more pronounced antiapoptotic effect on β cells.
The gene and protein expressions of MAFA, which is a marker of β-cell maturation, and MAFB, which is a marker of β-cell dedifferentiation, were significantly downregulated in the islets of rats with pre-DM following both A. muciniphila treatment and dietary intervention, particularly in those of the CD + AKK and HFD + AKK groups (p < 0.01) (Fig. 2F-H). These findings suggest that A. muciniphila treatment and dietary intervention can hinder the maturation of β cells and inhibit the dedifferentiation of β cells into α cells in rats with pre-DM. Additionally, the inhibitory effect of A. muciniphila on β-cell dedifferentiation is independent of diet.
Effects of dietary intervention andA. muciniphilatreatment on the intestinal flora and intestinal mucosal barrier in rats with pre-DM
After A. muciniphila treatment and dietary intervention for 5 weeks, DNA fragments extracted from the fecal samples of the rats in each group were sequenced with the Illumina platform. The Venn diagram plot of the intestinal flora illustrated that the number of specific ASVs in the HFD group was less than that in the control group. Dietary intervention increased the quantity of specific ASVs in the intestinal flora of rats with pre-DM, while A. muciniphila treatment had a minimal effect on the quantity of ASVs (Fig. 3A). Alpha diversity indices, such as Chao1 richness estimator, Faith’s PD, Shannon diversity index, and Observed species in the gut microbiome in the control group and each prediabetic group were analyzed (Fig. 3B). The beta diversity of the gut microbiome in each group was analyzed by PCoA and NMDS based on the weighted UniFrac distance and compared among the groups by PERMANOVA (Fig. 3C-E). These data suggest that dietary intervention can significantly improve the alpha diversity and beta diversity of the gut microbiome in rats with pre-DM (p < 0.05, p < 0.01 or p < 0.001), while A. muciniphila supplementation has no observable effects under the same dietary conditions.
At the phylum level, the dominant constituents of the gut microbiota in our rat model included Firmicutes (70.77%), Bacteroides (23.5%), and Proteobacteria (3.51%) (Fig. 4A). Dietary intervention significantly decreased the relative abundance of Firmicutes, Proteobacteria, and Deferribacteres in the intestinal flora of the HFD-induced prediabetic rats while increasing the relative abundance of Bacteroidetes and Elusimicrobia (p < 0.05, p < 0.01 or p < 0.001). A. muciniphila had little effect on the relative abundance of gut microbes in the rats with pre-DM (Fig. 4B). At the genus level, the predominant components of the gut microbiota in our rat model
were Ruminococcus (12.5%), Lactobacillus (6.42%), Oscillospira (5.04%), Prevotella (1.75%), Phascolarctobacterium (1.36%), and Blautia (1.27%) (Fig. 4C). We compared the differences in relative abundance of the top 20 genera in the intestinal flora of each group. Dietary intervention resulted in a significant increase in the relative abundance of Lactobacillus, Prevotella, and Turicibacter in the HFD-induced prediabetic rats, while the relative abundance of Oscillospira, Enterococcus, and Dorea decreased significantly (p < 0.05 or p < 0.01). A. muciniphila had a minimal effect on the relative abundance of the intestinal flora among the groups fed the same diets (Fig. 4D).
Histopathological analysis of the ileum revealed that the villi in the HFD group were notably atrophied, which was evident by the shortened and rounded morphologies. Moreover, the quantity of goblet cells in the ileum decreased significantly. Although the quantity of goblet cells in the HFD + AKK group decreased only slightly, a similar degree of villous atrophy was observed in comparison with the HFD group. In the ileal mucosa in the CD and CD + AKK groups, significantly less villi damage was observed, and the number of goblet cells increased significantly compared with those in the HFD group (Fig. 5A). Dietary intervention in the HFD-induced prediabetic rats significantly upregulated the protein expression of occludin (p < 0.05), which is an enzyme involved in the formation of tight junctions in the ileal mucosa (Fig. 5B).
Therefore, dietary intervention significantly improved the composition and diversity of the gut microbiome and repaired the intestinal barrier in the rats with pre-DM, while A. muciniphila supplementation alone had little effect.
A. muciniphila can reduce the levels of metaflammation in the circulation of rats with pre-DM
A. muciniphila treatment and dietary intervention both significantly reduced the serum levels of the proinflammatory cytokines LBP and IL-1β in rats with pre-DM (p < 0.01) (Fig. 6A, B). A. muciniphila treatment in conjunction with dietary intervention increased serum levels of the anti-inflammatory cytokine IL-10 in rats with pre-DM (p < 0.05) (Fig. 6C).
A. muciniphila can inhibit the TLR2 and TLR4 signaling pathways in the pancreatic islets of rats with pre-DM
Following A. muciniphila treatment and dietary intervention for 5 weeks, there was no significant difference in TLR2 gene expression in the islets of rats between groups (p > 0.05) (Fig. 6D). Both A. muciniphila supplementation and dietary intervention significantly downregulated the protein expression of TLR2 in the islets of HFD-induced prediabetic rats, while the effect of dietary intervention was more pronounced (p < 0.05 or p < 0.01) (Fig. 6H). Each treatment significantly downregulated the gene and protein expression of TLR4 in the islets of rats with pre-DM (p < 0.05 or p < 0.01) (Fig. 6E, H). A. muciniphila inhibited the expression of the inflammatory pathway TLR4/MyD88/NF-κB in the islets of rats with pre-DM (p < 0.05). A. muciniphila supplementation combined with dietary intervention resulted in a more pronounced inhibitory effect on the TLR4 inflammatory pathway (p < 0.01) (Fig. 6E-H). These findings suggest that the gut microbiota may contribute to the regulation of β-cell apoptosis, differentiation, and function in rats with pre-DM through inflammatory pathways mediated by TLR2 and TLR4.
{kind=link}