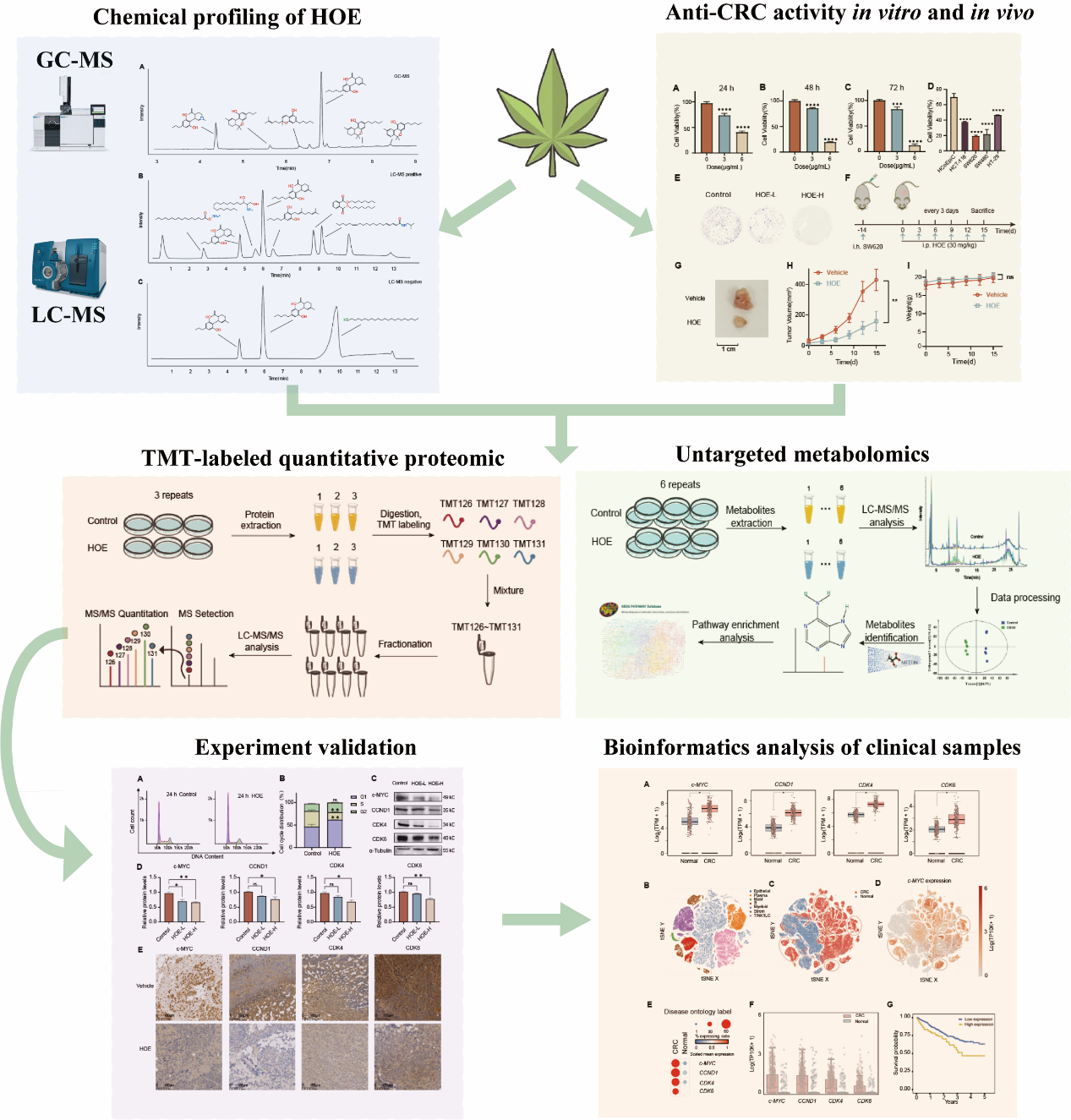
3.1. Chemical profiling of HOE
In this current investigation, we employed a comprehensive analytical approach utilizing UPLC-MS coupled with GC-MS to delineate the specific components within HOE. The mass spectra of both GC (Fig. 1A) and UPLC (Fig. 1B-C) are presented in Fig. 1. Through a meticulous comparison of mass spectrum data and consideration of physicochemical properties of known compounds, we successfully identified 13 compounds through UPLC-MS and 6 compounds through GC-MS. Ultimately, a total of 16 representative compounds, including cannabidiol, cannabidivarin, cannabinol, among others, were tentatively identified. This robust identification establishes a foundational framework for subsequent research endeavors.
3.2. HOE inhibits CRC cell proliferation in vitro and in vivo
To investigate the impact of HOE on CRC cell proliferation, SW620 cells were treated with 3 and 6 µg/mL of HOE for 24, 48 and 72 h. The results showed that the cell viability of SW620 decreased in a time and dose-dependent manner (Fig. 2A-C). In comparison to the normal colon primary cell line, HCoEpic, HOE exhibited a greater inhibitory effect in various CRC cells such as HCT-116, SW620, SW480, and HT-29 (Fig. 2D), indicating that HOE has a general inhibitory effect on CRC cell growth while causing minimal damage to normal cells. As depicted in Fig. 2E, HOE suppressed colony formation in SW620 cells after two weeks of treatment, with higher concentrations yielding higher inhibition rates.
To assess the potential effects of HOE on CRC in vivo, a xenograft model using the SW620 cell line was established in nude mice(Fig. 2F). As shown in Fig. 2G, HOE induced a significant reduction in tumor volume, the body weight and tumor volume of mice were measured every three days, and the results demonstrated that HOE could inhibit the tumor growth while not significantly affecting the mice’s weight (Fig. 2H-I). H&E staining revealed that the number of tumor cells in the model group is greater than in the HOE treatment group (Fig. 2J-K). Additionally, the protein levels of the proliferation marker Ki-67 were determined by IHC staining (Fig. 2L-M), which showed that the treatment with HOE decreased Ki-67 expression. These findings indicate that HOE suppresses tumor cell proliferation in the CRC xenograft model. Overall, these observations demonstrate that HOE can inhibit CRC proliferation in vitro and in vivo with minimal side effects.
3.3. Quantitative proteomics-coupled with bioinformatics displays the landscape of HOE-regulated protein networks
To validate and reveal the potential mechanism of HOE in treating of CRC, the quantitative proteomics was conducted in comparison of the HOE treatment group and the vehicle group. The protein was initially extracted from the cells and subsequently hydrolyzed into peptides, which were subsequently labeled with TMT agents. Following mixing and fractionation, the resultant samples were subsequently introduced into the UHPLC-MS/MS system (Fig. 3A). A total of 5853 proteins, including 134 up-regulated proteins and 139 down-regulated proteins, were identified as the differentially expressed proteins (DEPs). The volcano plot was presented based on the threshold value of |Fold change| ≥ 1.2 and p-value < 0.05. Figure 3B encapsulates the dysregulated proteins inclusive of CDK4, PLK1, CCNB2, c-MYC, exhibiting downregulation, and APOB, GPAT3, depicting upregulation. Accordingly, the protein-protein interaction network of the DEPs was generated, with c-MYC exhibiting the highest degree value as one of the significant hubs of the network (Fig. 3C), indicating the crucial role of c-MYC in the anti-tumor effects of HOE. To further investigate the potential mechanism, KEGG functional enrichment results of these dysregulated proteins are presented (Fig. 3D), with the top 15 signaling pathways enriched including cell cycle, DNA replication, pyrimidine metabolism, purine metabolism, etc.. And cluster and enrichment analysis was conducted with Metascape. The DEPs were colored by cluster ID, where nodes that share the same cluster ID are typically close to each other and belong to the same pathway, which including cell cycle, DNA metabolic process, mitotic cell cycle, etc. (Fig. 3E). Then, we used DAVID to perform the GO analysis to reveal the functions of DEPs. The significant GO terms and major proteins were shown in the chord plot, which demonstrating that the functions of DEPs are tightly linked to the cell cycle pathway. Overall, the results of the quantitative proteomic confirmed that HOE could exhibit anti-tumor effects by affecting the cell cycle pathway.
3.4. Metabolomics reveals the profiling of HOE-regulated metabolites
Metabolomics analysis was performed to explore the metabolic alterations upon HOE (3 ug/mL) treatment. In order to delineate differential metabolite profiles between the control and HOE groups, OPLS-DA models were meticulously constructed. In this context, the established OPLS-DA models (Fig. 4A-B) exhibited a commendable fit and impressive predictability.
Next, for further refinement in the selection of differential metabolites, the volcano plot was presented to depict differential metabolites after HOE treatment in the positive (C) and negative (D) modes, respectively(Fig. 4C-D). Based on the threshold value of fold change (≤ 0.83 or ≥ 1.2) and p-value (< 0.05), 1056 differential ions in positive mode and 1915 differential ions in negative mode were acquired. 53 metabolites showing significant differences in the HOE treatment group were identified, which were markedly differentiable from the control group, whose relative levels between the two groups were significantly different demonstrated in Heatmap determined by LC-MS, normalized by Z-score (Fig. 4E). Of significant note, a marked decrease was observed in the levels of crucial metabolites, including ATP (FC = 0.28, p = 0.001), GTP (FC = 0.22, p = 0.003), adenine (FC = 0.25, p = 0.001), and Guanine (FC = 0.72, p = 0.015), within the purine metabolic pathway. Conversely, a significant increase was observed in the concentration of GMP (FC = 1.34, p = 0.004), xanthosine (FC = 5.04, p = 0.001), and xanthine (FC = 4.36, p = 0.001).
Functional enrichment analysis, grounded in the altered metabolites, revealed the top 11 distinctive metabolic pathways differentiating the two groups, as depicted in Fig. 4F. These pathways encompassed purine metabolism, pyrimidine metabolism, nucleotide sugars metabolism, among others. In summary, the pathways of purine and pyrimidine metabolism underwent significant alterations following HOE treatment, primarily exhibiting a discernible decreasing trend.
Given the limitations of functional enrichment analysis relying on sparse biochemical knowledge annotations [21], we sought to delve deeper into the altered metabolites from the perspective of chemical structure. For this purpose, we employed ChemRICH analysis, a statistical enrichment approach that focuses on chemical similarity to cluster metabolites with similar structures. ChemRICH statistics revealed significant differences in purine nucleosides, specifically adenine and guanine nucleotides, and pyrimidine nucleosides (Fig. 4G). Simultaneously, a Tanimoto chemical similarity clustering tree was constructed to visually represent the clustering of metabolites with distinct structures, as illustrated in Fig. 4H. Figure 4I illustrates the specific relative levels of metabolites associated with purine metabolism, all of which exhibited significant differences between the HOE-treated and control groups.
In summary, our findings suggest that HOE disrupted the purine and pyrimidine metabolism in CRC cells. As these endogenous substances are essential for cell cycle progression [22], the metabolome results are consistent with the proteome. This comprehensive analysis, combining functional enrichment and chemical structure perspectives, enhances our understanding of the intricate metabolic alterations induced by HOE treatment.
3.5. HOE arrests cells in the G1 phase by inhibiting c-MYC
Based on the result of integrated proteomics and metabolomics method, we hypothesized that HOE functioned as an anti-CRC agent by arresting cell cycle via c-MYC, leading to the disruption of purine and pyrimidine metabolism. Flow cytometric analysis showed that G1-phase cells increased from 46.5–61.9% after treatment of HOE on SW620 cells for 24 h (Fig. 5A-B), which indicated that HOE arrested the cell cycle at the G1 phase in SW620 cells. To further confirm the mechanism of HOE on the cell cycle of the SW620 cell line, we examined the protein expression of c-MYC and other cycle-related proteins by western blotting (Fig. 5C-D). The results showed that HOE dose-dependently decreased the protein expression of c-MYC, CCND1, CDK4, and CDK6. Using the xenograft mouse model, we measured the levels of these four proteins by means of immunohistochemistry staining, whose results were consistent with the cellular level demonstrating exposure to HOE occurred with varied degrees of decrease (Fig. 5E-F). Therefore, HOE arrested the cell cycle of SW620 by modulating the level of c-MYC to realize the inhibition of carcinogenesis and progression of CRC.
3.6. Bioinformatics analysis unraveled specific gene expression differences and heterogeneity in CRC
To assess the potential clinical applicability of HOE, we comprehensively evaluated the expression profiles of the four critical genes, i.e., c-MYC, CCND1, CDK4, and CDK6, in clinical samples of CRC and adjacent normal tissues. By utilizing both bulk RNA-seq and single-cell RNA-seq data, we aimed to investigate whether the four genes displayed heightened expression levels within the tumor microenvironment. Our analysis of The Cancer Genome Atlas (TCGA) dataset, as depicted in Fig. 6A, revealed a statistically significant upregulation of these genes within CRC tissue, suggesting their pivotal involvement in the etiology and progression of CRC. Moreover, employing single-cell RNA-seq analysis, we further embarked on an exhaustive quest to elucidate the cellular heterogeneity present in 62 CRC and 36 adjacent normal colon specimens. Our meticulous efforts led to the identification of distinct cell types, including epithelial cells (n = 16,135), B cells (n = 16,134), master cells (n = 3,872), myeloid cells (n = 16,134), plasma cells (n = 16,134), stromal cells (n = 15,457), and T/NK/ILC cells (n = 16,134), as depicted in Fig. 6B. Considering that CRC lesions predominantly manifest in colorectal epithelial cells, we conducted an in-depth clustering analysis of epithelial cells in both CRC and normal contexts. Through the utilization of t-SNE visualization, we successfully demarcated the epithelial cell subcluster into two distinctive clusters (Fig. 6C). Then we focused on investigating c-MYC expression patterns within the epithelial cell subcluster, revealing notable disparities among individual cells. Specifically, upregulation of c-MYC was observed in the CRC cluster (Fig. 6D). To assess gene expression heterogeneity comprehensively, we constructed a dot plot (Fig. 6E) that captured the ratio of cells expressing specific genes to the total population while evaluating scaled mean expression levels across different labels. Quantitative analysis of c-MYC, CCND1, CDK4, and CDK6 expression in CRC and normal cells confirmed pronounced heterogeneity (Fig. 6F). Remarkably, patients were categorized into two groups based on the expression levels of these genes. Survival time, representing the duration without disease progression during and after treatment, revealed significant differences between the groups (Fig. 6G), highlighting the clinical relevance of these genes. Our bioinformatic analysis provided valuable insights into gene expression differences between CRC and adjacent normal tissues, emphasizing the therapeutic potential of targeting these four genes and the promising clinical applications of HOE.
{kind=link}