Viral infection up-regulated the expression of PARP14 in macrophages.
In the previous study, we observed that most of the PARP members could be up-regulated upon both VSV and HSV infection, such as Parp3, Parp8, Parp9, Parp10, Parp11, Parp12, Parp13, Parp14, and so on, which correlated with the expression of Ifnb1, Isg15, and Cxcl10 (Fig. 1A). As shown in Fig. 1B, we found that Parp14 was also up-regulated in the lung tissue of Rhesus macaques infected by SARS-CoV-2 on day 3 and day 7 through re-analyzing the publicly published data (GSE158297). These data indicated that Parp14 may play a role in the anti-viral process of the host. Then, we search for the expression pattern of PARP14 in the Human Protein Atlas database (https://www.proteinatlas.org/)26. Data showed that Parp14 was widely expressed in different human tissues; the spleen, tonsil, liver, liver, lung, and so on were the top 10 tissues (Fig. 1C). It was also worth noting that the Parp14 gene was highly expressed in the human immune cells, such as B-cells, Macrophages, Monocytes, NK cells, and T cells (Fig. 1D), suggesting its function may relate to the host's immune system. To validate these predictions, we examined the expression of Parp14 at different immune cells with various viruses infection (Fig. 1E-G). The results shown in Fig. 1E show that MHV, HSV, and VSV infection could significantly induce the expression of Parp14 compared to the mock group in BMDMs, except for the SeV infection. In the PMs and RAW 264.7 cells, all of these viruses up-regulated the expression level of Parp14 significantly (Fig. 1F-G). Together, these results demonstrated that virus infection could up-regulate the expression of Parp14 in macrophages.
Depletion of PARP14 enhances the expression of Il1b and Il6 in macrophages.
Then, we generated the Parp14 knock-out (KO) mice and macrophages to explore the potential roles of Parp14 in the host immune system. Unexpectedly, the transcription of Ifnb1 from the WT and KO groups had no significant difference with the infection of HSV or VSV for 6 hours (Fig. 2A-B). However, the expression of Il1b and Il6 were significantly up-regulated in Parp14 KO PMs upon the infection of HSV and VSV, respectively (Fig. 2A-B). Consistent with the results of PMs, the expression of Ifnb1 in the Parp14 KO was weaker than WT BMDMs but did not meet the significant difference upon the infection of HSV, SeV, and VSV (Fig. 2C). In contrast, the Il6 had a more robust expression in Parp14 KO BMDMs upon viral infection.
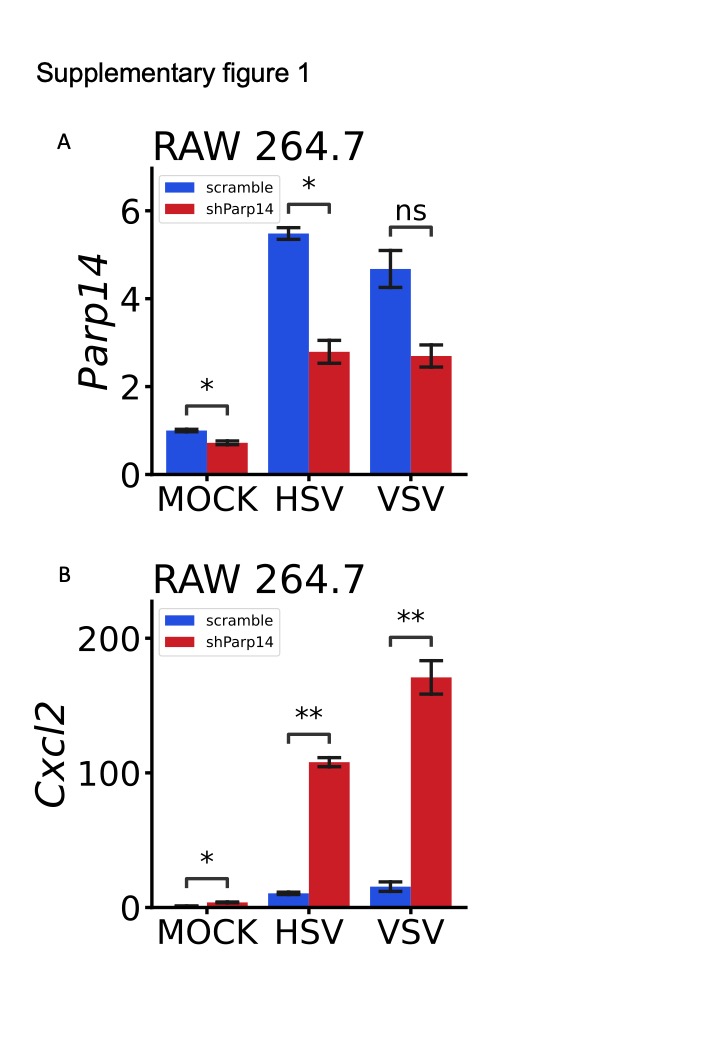
To validate these results, we used the shRNA of Parp14 to generate the Parp14 knock-down (KD) RAW 264.7 cells. The primer sequences of shRNA for Parp14 are shown in Table S2. The effect of shRNA to inhibit the expression of Parp14 was shown in Fig S1A, which could significantly down-regulate the expression of Parp14 at RAW 264.7 cells with or without virus treatment. Consistent with these results from BMDMs and PMs, the expression of Il1b and Il6 rather than Ifnb1 were significantly up-regulated in KD RAW 264.7 cells upon HSV and VSV treatment compared to the WT RAW 264.7 cells. We also found that the expression of Cxcl2 was up-regulated considerably upon HSV and VSV treatment (Fig S1B). Overall, these results demonstrated that KO or KD Parp14 in the macrophages almost had no significant influence on Ifnb1 but significantly elicited the expression of Il1b and Il6, which indicated that Parp14 may play roles in the host's inflammatory response upon stimulations.
KO of PARP14 exhibits a more robust inflammatory response upon virus infection in macrophages.
To depict the influence of Parp14 KO on host cells at the whole transcriptome level, we conducted the bulk RNA-seq on Parp14 KO and WT BMDMs upon HSV and VSV infection. As shown in Fig. 3A, some genes were up-regulated or down-regulated regardless of HSV or VSV infection. For example, Arg1, Csta2, Csf3, Il1b, Pf4, and S100a8 genes were induced more highly upon both HSV and VSV infection in the KO group compared to the WT group, while Tlr9, Wdfy1, and Itpr1 genes in KO group were lower than WT BMDMs. Figure 3B shows that Parp14 and some interferon genes were significantly induced in WT BMDMs upon HSV and VSV treatment. At the same time, some inflammatory cytokines, such as Il1a, Il1b, Pf4, Cxcl2, and Tnfsf9, had a higher expression level in the KO BMDMs group. Then, we performed the KEGG pathway enrichment and GO annotation analysis for the DEGs between the KO and WT BMDMs upon HSV and VSV treatment, respectively (Fig. 3C-F). As shown in Fig. 3C and Fig. 3E, the up-regulated genes were enriched into these pathways, which included Hematopoietic cell lineage, Rheumatoid arthritis, Cytokine-cytokine receptor interaction, Th17 cell differentiation, Th1 and Th2 cell differentiation, T cell receptor signaling pathway and inflammatory bowel disease and so on regardless of HSV or VSV infection. The GO annotation results of up-regulated DEGs upon HSV and VSV treatment were shown in Fig. 3D and Fig. 3F: immune system process, adaptive immune response, MHC class II protein complex, positive regulation of T cell activation, cellular response to interferon-pathway and neutrophil chemotaxis.
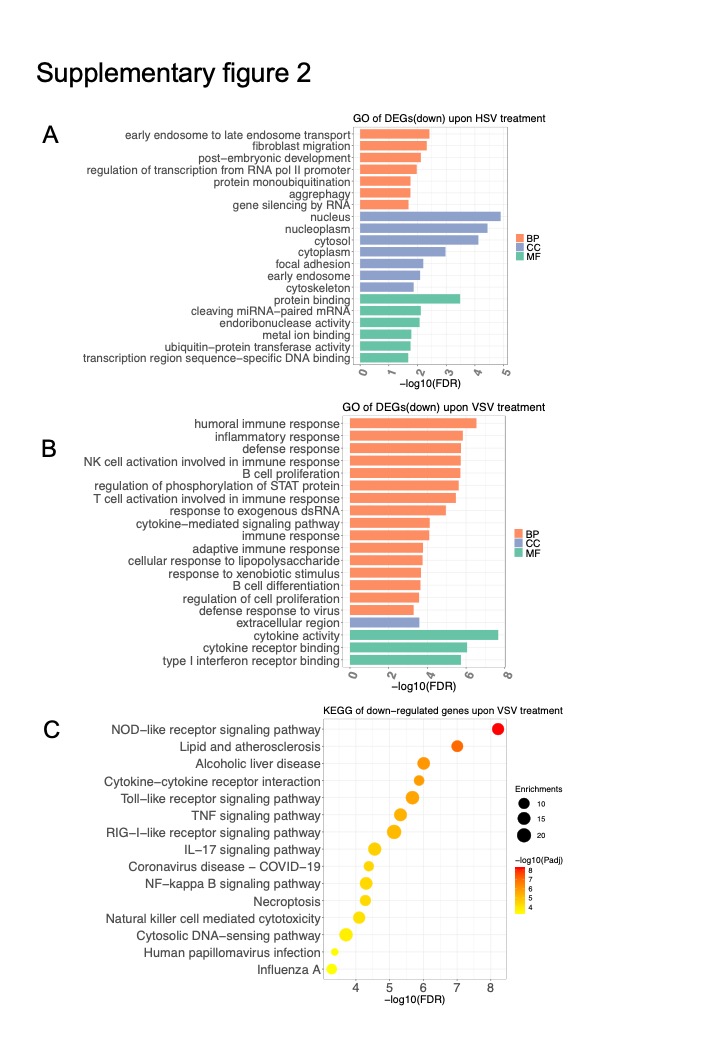
The GO enrichment analysis results of down-regulated DEGs in the KO BMDMs were shown in Fig S2A-B, which contained the protein monoubiquitination, gene silencing by RNA, regulation of phosphorylation of STAT proteins, and immune response. The KEGG enrichment analysis of significantly down-regulated genes in the KO group upon VSV treatment included some innate immune-related responses, such as NOD-like receptor signaling pathway, Toll-like receptor signaling pathway, RIG-I-like receptor signaling pathway, IL-17 signaling pathway (Fig S3C). Then, we focused on the DEGs between Parp14 KO and WT upon VSV infection by conducting a PPI network analysis. As shown in Fig. 3G-H, some up-regulated DEGs were enriched in the inflammatory response, while the down-regulated DEGs were enriched in the innate immune response. Together, these results suggested that KO of Parp14 impaired the innate immune response but enhanced the inflammatory response of the host.
Knockout of Parp14 increases the susceptibility to LPS challenge.
Because the Parp14 KO macrophages exhibited a more robust inflammatory response, we examined whether there was a distinguished response to the LPS challenge between the KO and WT groups. As shown in Fig. 4A, LPS treatment could not generate a significant difference in the expression of Ifnb1 but significantly induced the transcription of Il6 and Il1b, consistent with the treatment of HSV and VSV. Moreover, LPS treatment also induced a higher expression level of inflammatory cytokines rather than the expression of Ifnb1 (Fig. 4B). To further validate the anti-inflammatory effect of Parp14, we injected the Parp14 KO and WT mice with VSV and LPS, respectively. As expected, the survival time of KO and WT mice had no significant difference upon VSV infection, consistent with these results at the cell level. However, the survival time of WT mice was significantly longer than the Parp14 KO mice. Then, we examined the expression of some inflammatory cytokines between the KO and WT mice. As shown in Fig. 4E, upon LPS injection 10 hours, the Tnf, Il6, and Il1b genes were significantly higher than the WT mice, which may be the main reason for the death of Parp14 KO mice. Together, these data demonstrated that Parp14 played a stronger anti-inflammatory function than its anti-viral effect.
PARP14 inhibits the inflammatory response through the NKFB pathway
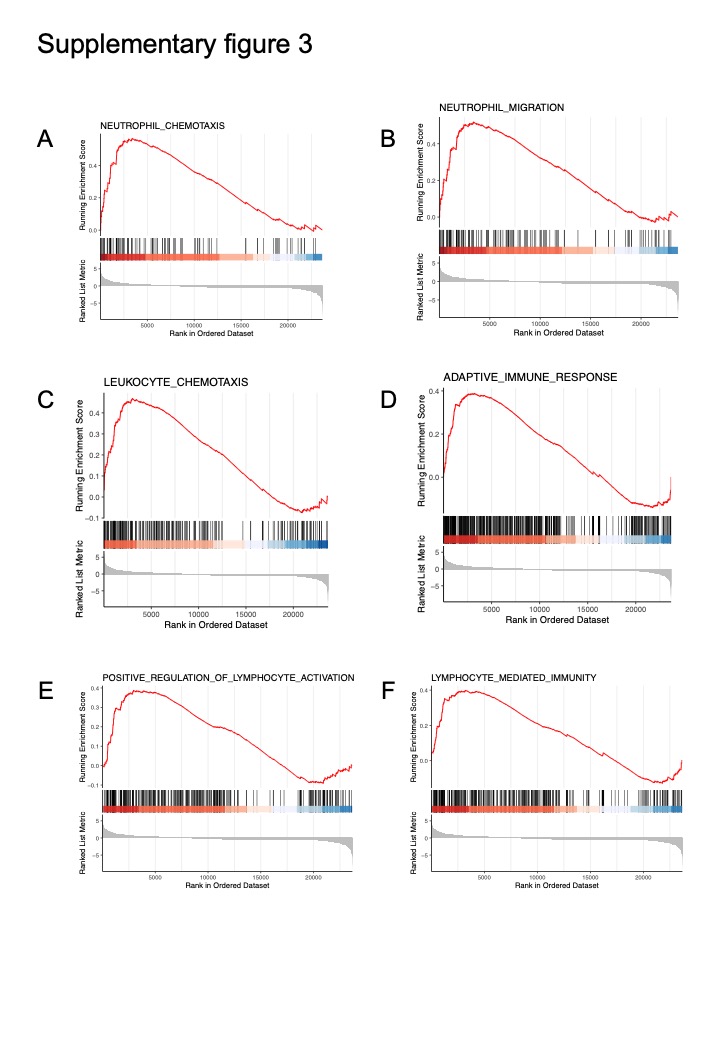
To further explore the molecular mechanism by which Parp14 plays the anti-inflammatory role, we conducted the RNA-seq on the PMs derived from the KO and WT mice and then treated them with LPS for 6 hours. The DEGs shown in Fig. 5A, Arg1, Cxcl13, Gzma, Gzmc, Il1b, and Cxcr3 were significantly up-regulated, while Fgg, Fga, Fgb, Apoa1, Apoh, and Alb were down-regulated considerably in KO compared to WT group. KEGG pathway enrichment analysis was conducted on the up-regulated DEGs in the KO group, and the results were shown in Fig. 5B, which contained adaptive immune response, leukocyte cell-cell adhesion, regulation of T cell activation, leukocyte migration, lymphocyte differentiation and lymphocyte-mediated immunity and so on. Then, we conducted the GSEA between KO and WT PMs (Fig. 5C-D). The top enriched terms included regulation of innate immune response, positive regulation of inflammatory response, neutrophil chemotaxis, neutrophil migration, leukocyte chemotaxis, adaptive immune response, positive regulation of lymphocyte activation, and lymphocyte-mediated immunity (Fig S3A-F).
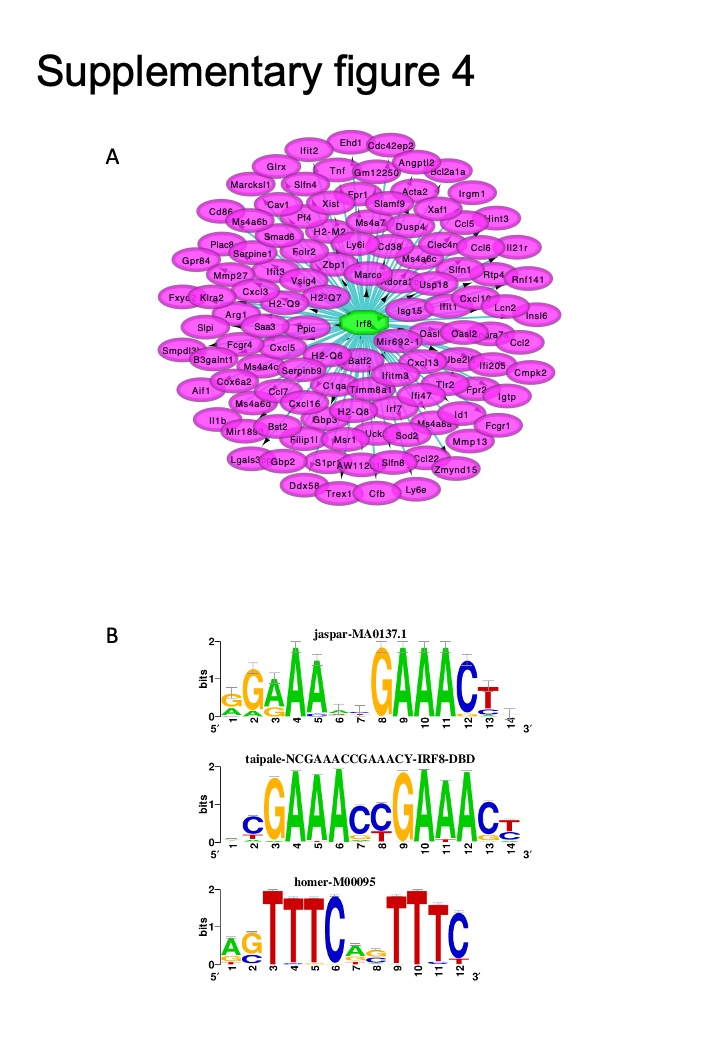
Then, we used the up-regulated DEGs in KO PMs to perform the TFEA with Cytoscape software and iRegulon plug-in. As shown in Fig. 5E, NF-κB1 was the top enriched transcription factor (TF) that regulated most of these up-regulated genes. The predicted DNA motif sequences bound by the NF-κB1 are shown in Fig. 5F. The second enriched TF was IRF8; the predicted binding motifs are shown in Fig S4A-B. These results indicated that Parp14 may regulate the expression of inflammatory cytokines through the NF-κB or IRF8. Thus, we used the inhibitor of NF-ΚB, JSH23, to pre-treat KO and WT PMs before LPS treatment. As shown in Fig. 5G, JSH23 almost eliminated the difference of Il6 and Il1b between WT and KO groups, which demonstrated that Parp14 inhibited the expression of some inflammatory cytokines through the NF-κB pathway.
{kind=link}
{kind=link}
{kind=link}
{kind=link}