Constructing of transcriptome profiles of Euphorbia kansui after MeJA treatment.
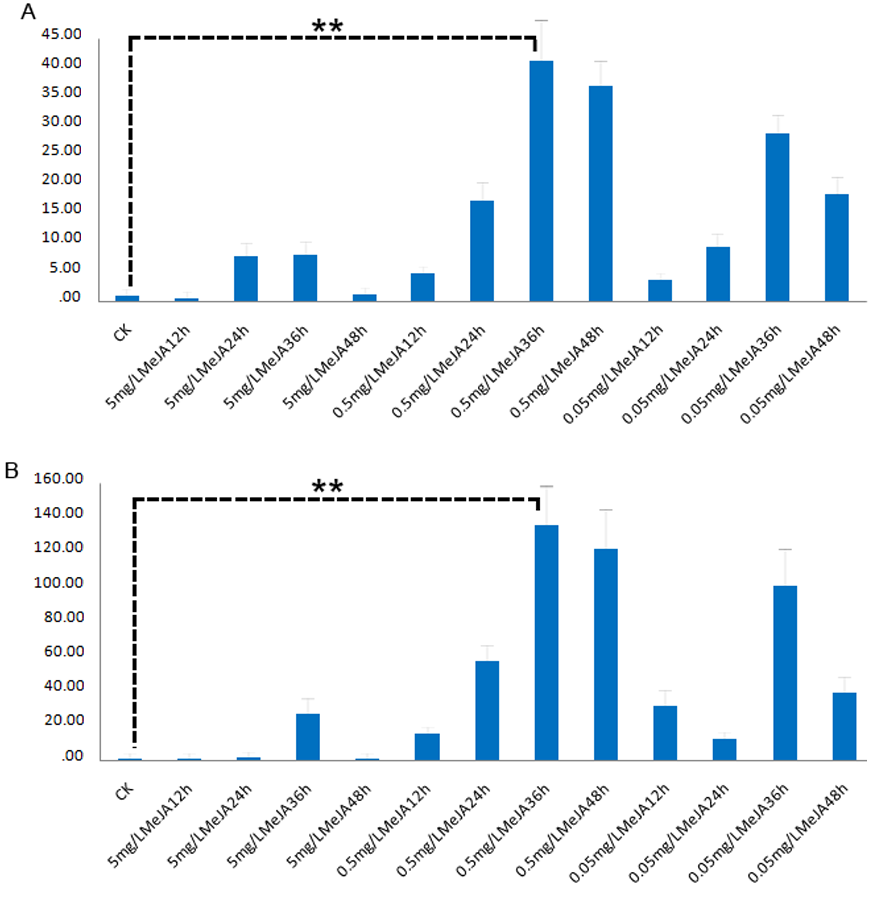
Exogenous MeJA treatments on Euphorbia kansui (EK) seedlings were conducted in three different concentrations, including 200 µM, 20 µM and 2 µM of MeJA. Casbene and AACT were chosen to evaluate the effects of different concentration of MeJA on causing changes of gene expression level in EK. In all spraying experiments, the expression levels of casbene and AACT increased at 24 h, 36 h after MeJA treatment (hat), but did not exhibite significant change at 12 hat, then slightly reduced at 48 hat (Figure S1). However, only 20 µM of exogenous MeJA treatment resulted in significant increases in expression levels of both genes at 24 hat, 36 hat, and significantly decreased at 48 hat (Figure S1; P < 0.05). Thus, 20 µM MeJA spraying experiments at 0 (CK), 24 (T1), 36 (T2), 48 (T3) hat were chosen for transcriptome sequencing.
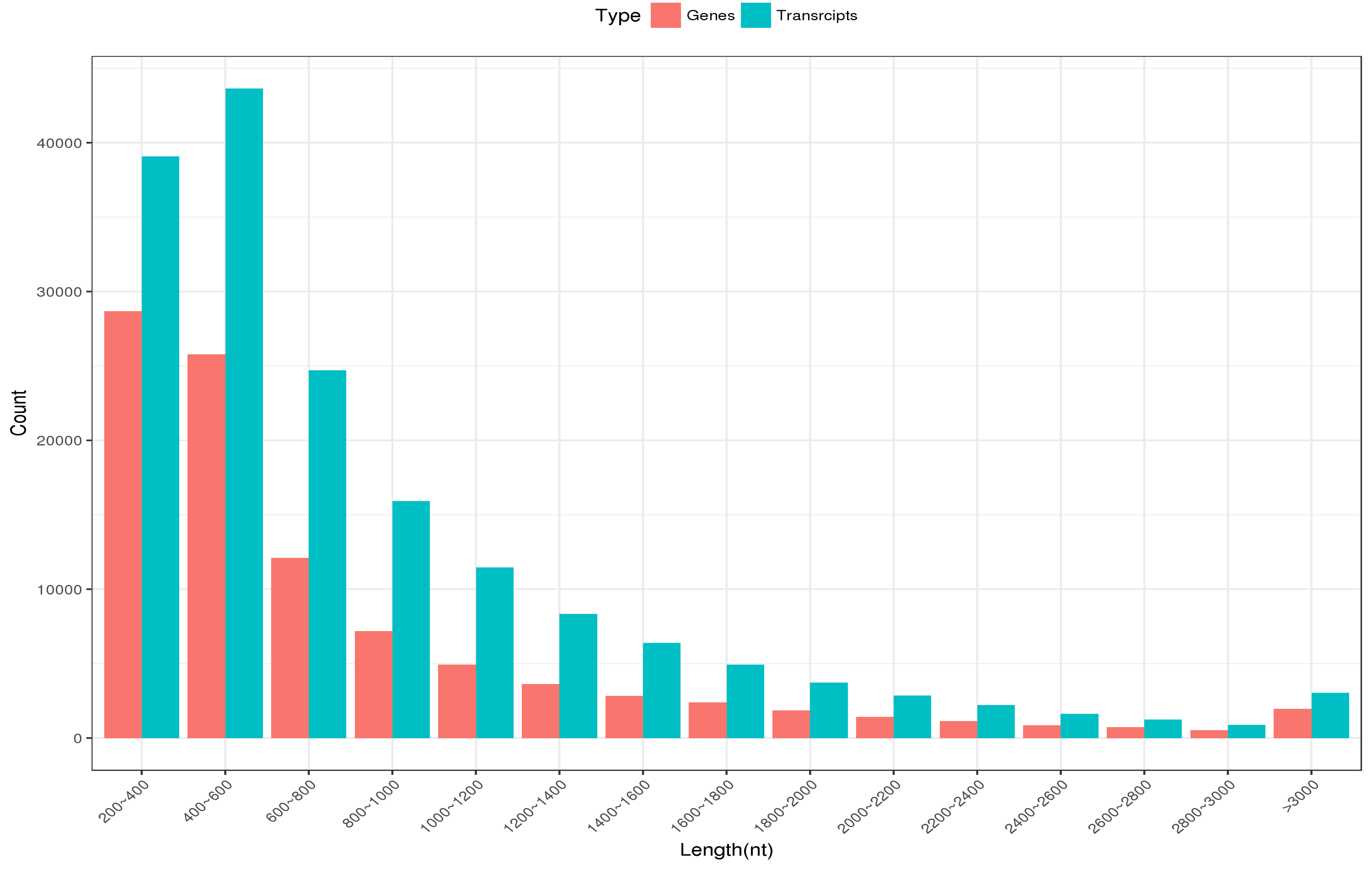
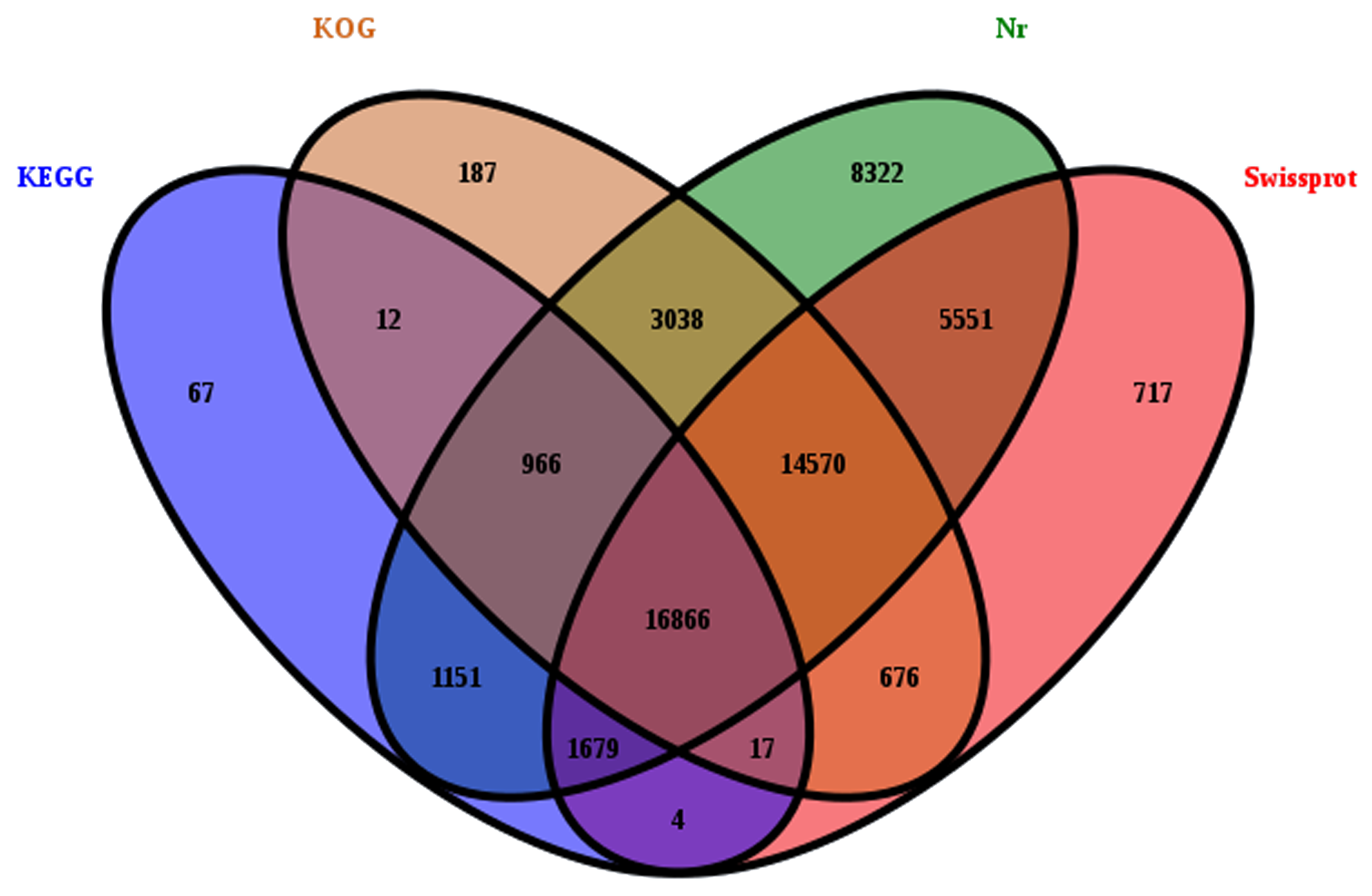
For transcriptome profiles of EK following MeJA treatment, three biological replicates from each experimental group were collected for sequencing. Through raw reads filtering and trinity assembly, we obtained 96,040 unigenes with an average length of 812 bp (Table 1, Figure S2). Then, we analyzed the function of these unigenes using numerous function annotation databases, including Nr, Swiss-Prot, KEGG and KOG databases (Figure S3). Typically, among all unigenes, 52,142 unigenes showed significant similarities to the known proteins in the Nr database, 40,079 had BLAST hits in the Swiss-Prot database, 20,761 in the KEGG database and 36,331 in KOG database (Figure S3). In the Nr and Swiss-Prot databases, most of the matched sequences (69.87% in Nr and 67.69% in Swiss-Prot) had E-values between 0 and 1 ×10− 20, while the rest 30.13% in Nr and 32.31% in Swiss-Prot had E-values from 1 × 10− 20 to 1 × 10− 5 (Fig. 2A, B). Homologies among different species are illustrated in Fig. 2C, where out of the matched 52,142 unigenes in Nr, 17,511 (33.58%) were matched to Jatropha curcas followed by Populus euphratica (3107; 5.96%) and Theobroma cacao (2661; 5.10%), suggesting that E. kansui was genetically more related to Jatropha curcas.
Table 1
| Type | Amount | GC% | N50 | Min Length | Mean Length | Mac Length | Assembled Bases |
| Genes | 96040 | 39.1749 | 1066 | 300 | 812 | 12511 | 78037302 |
| Transcripts | 170087 | 38.8688 | 1116 | 300 | 865 | 12511 | 147240952 |
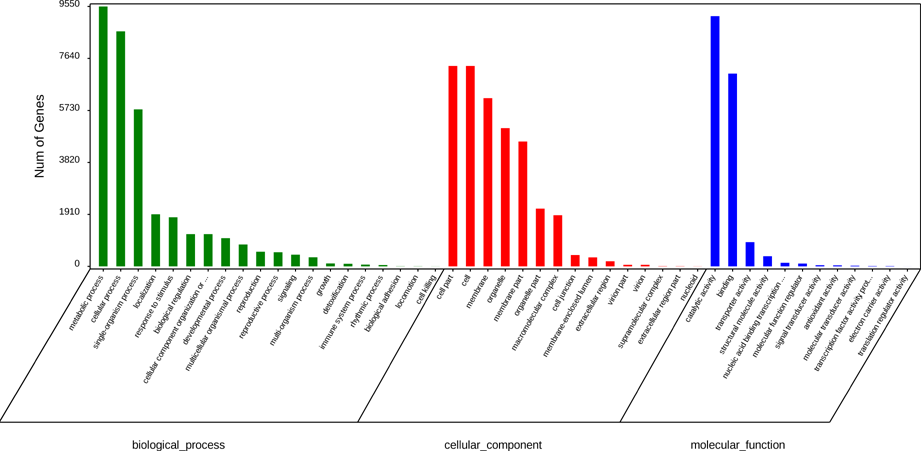
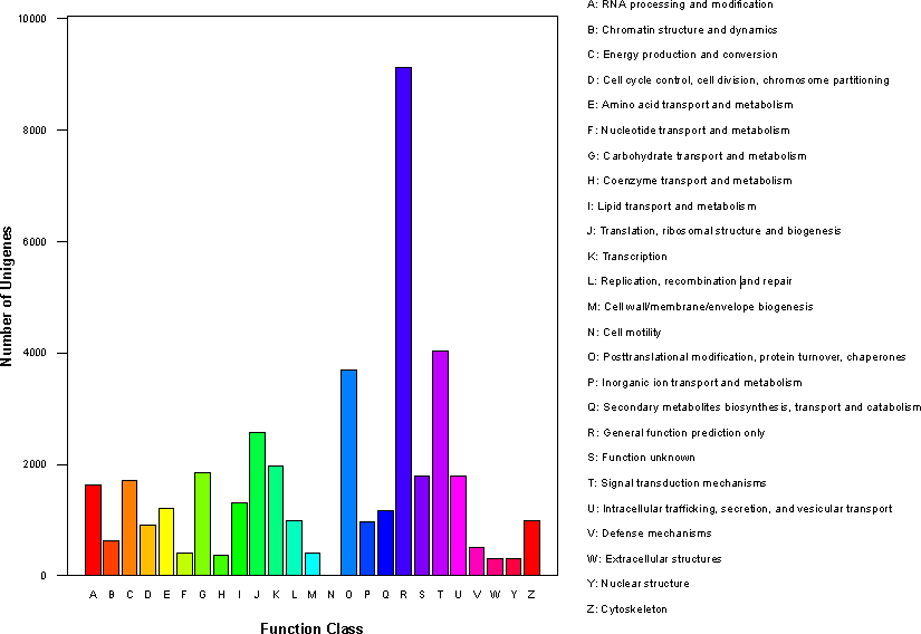
Subsequently, we annotated the 52,142 unigenes using GO, KEGG and KOG databases (Figure S4, S5). The results showed that GO assignments to the biological processes were the majority, followed by molecular function and cellular component (Figure S4; Additional file 2: Table S2). Meanwhile, the results of KOG showed that 36,331 unigenes were assigned to the KOG classifications (Figure S5; Table S3). Of note, both KEGG and KOG annotation showed that the clusters of secondary metabolites biosynthesis, lipid transport and metabolism, Terpenoid backbone biosynthesis, Flavonoid biosynthesis and Sesquiterpenoid and triterpenoid biosynthesis were the major groups, showing the ability of EK in producing secondary metabolites represented by terpenoids (Figure S5; Additional file 3: Table S3; Additional file 4: Table S4).
MeJA treatment caused dramatic changes in transcription pattern of Euphorbia kansui.
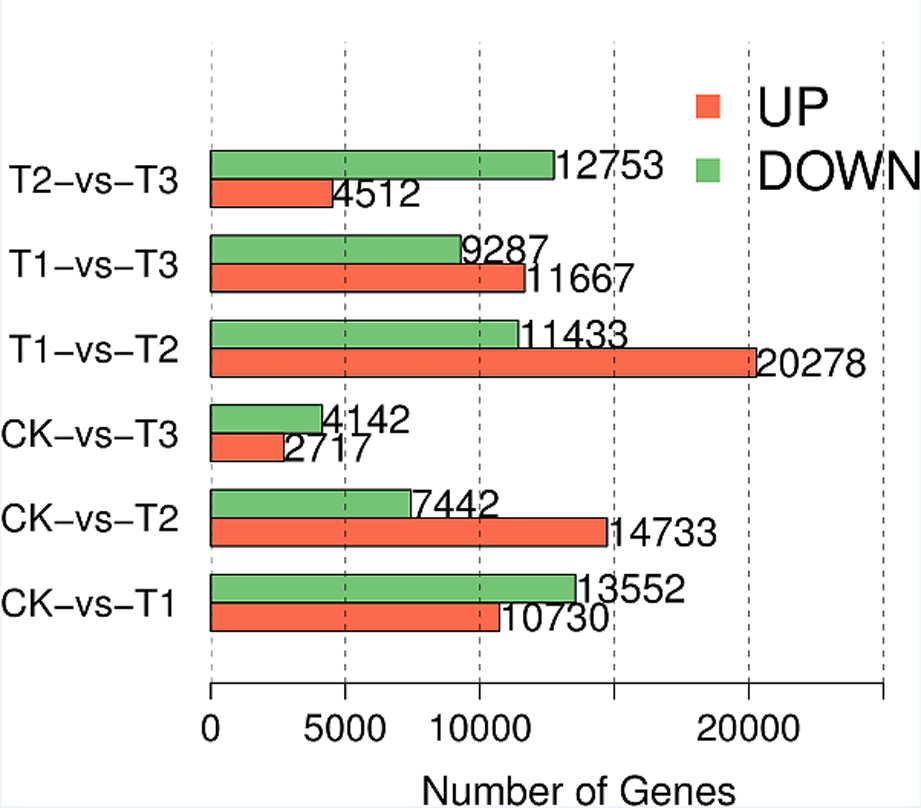
To investigate the variations in transcription pattern of EK following MeJA treatment at different time points, we performed principal component analysis (PCA) on all transcriptome profiles. As shown in Fig. 3A, the top 2 principal component were used to construct the PCA plots (Fig. 3A). The PCA plots showed that all biological replicates from the same experimental group were clustered together, and clear separated with other groups, suggesting that MeJA treatment caused significant changes in the transcription pattern of EK (Fig. 3A). The principal component 1 (PC1) explained 59.5% variations in transcription pattern of all samples, while these variations were mainly caused by MeJA treatment (Fig. 3A), supporting the effects of MeJA in altering transcription pattern of EK. Typically, the distance between TI and CK was larger than that between T2/T3 and CK, suggesting that exogenous application of MeJA mainly caused transcription variations in EK at 24 hat. Then, we analyzed the differentially expressed genes (DEGs) in each comparison using fold change ≥ 2 and a p (P-Value) < 0.05 (Fig. 3B-C; Figure S6). Totally, we identified 12,106 upregulated and 19,952 downregulated DEGs T1 vs. CK comparison, and 5,330 upregulated and 9,953 downregulated DEGs in T2 vs. CK, 4,281 upregulated and 8,185 downregulated DEGs in T3 vs. CK, respectively (Fig. 3B-C; Figure S6). Of note, the number of DEGs in T1 vs. CK was more than that in other two pairwise comparisons (Fig. 3B-C; Figure S6), supporting the phenomenon that exogenous application of MeJA mainly caused transcription variations in EK at 24 hat. Additionally, there are also plenty of DEGs in T1 vs. T2 and T1 vs. T3 comparisons, as shown in (Figure S6).
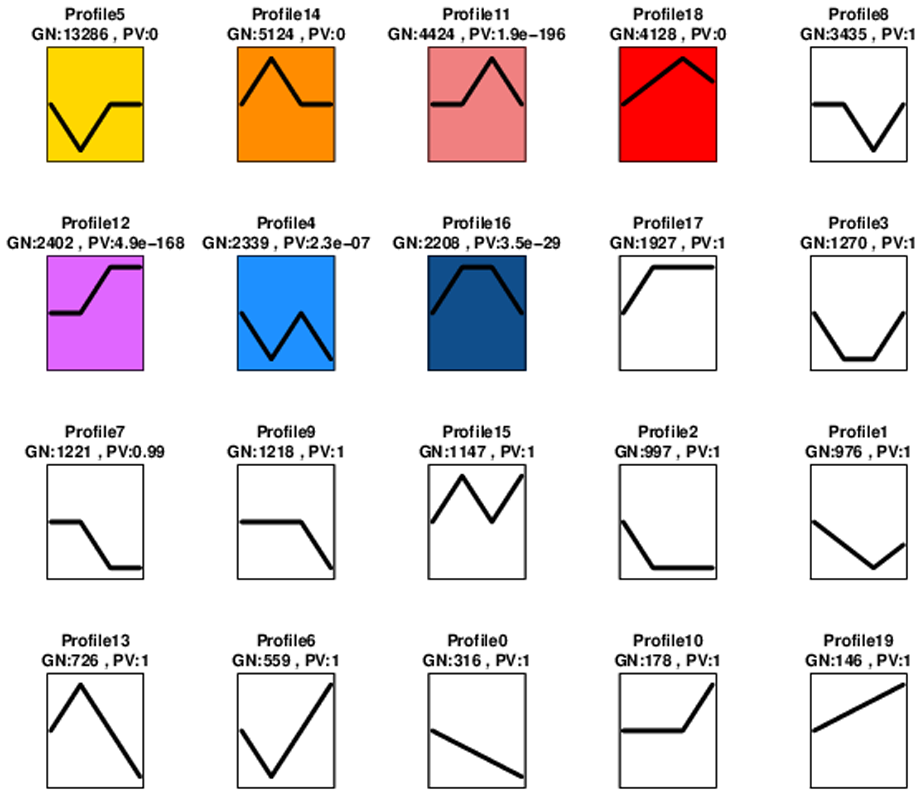
Subsequently, the Short Timeseries Expression Miner (STEM) cluster was used to directly compare the expression patterns of these DEGs in response to MeJA treatment of 0 h (CK), 24 hat (T1), 36 h (T2) and 48 h (T3). Seven significant gene profiles (P-Value ≤ 0.01): STEM profile 5 (13,286 genes), profile 14 (5,124 genes), profile 11 (4,424 genes), profile 18 (4,128 genes), profile 12 (2,402 genes), profile 4 (2,339 genes) and profile 16 (2,208 genes) were identified as significant changes (Figure S7). It could be found that in profile 11, 18 and 12, after MeJA treatment 36 hat, the gene expression levels reached the peaks, after MeJA treatment 48 hat, gene expression decreased in profile 11 and 18 and kept unchanged in profile 12 (Figure S7). While in profile 14 and 16, gene expression levels got to the highest at MeJA treatment 24 hat, and then decreased at treatment 36 hat and 48 hat, respectively (Figure S7). In profile 4 and 5, gene expression levels get to the lowest at MeJA treatment 24 hat, and then got back to that before MeJA treatment at MeJA 36 hat (Figure S7). Taken together, these results suggested that exogenous MeJA application dramatically altered the transcription pattern in EK, especially at 24 hat.
MeJA treatment altered the expression of genes with function relevant to terpenoid biosynthesis in Euphorbia kansui.
To investigate the function of DEGs associated with MeJA treatment at different time points in EK, we performed GO enrichment analysis on the DEGs in T1 vs. CK, T2 vs. CK and T3 vs. CK comparisons. In “Biological Process” between T1 and CK, we found the DEGs were mainly enriched in plastid organization, cell wall organization or biogenesis, shoot system morphogenesis, leaf morphogenesis, external encapsulating structure organization, reactive oxygen species metabolic process, carbohydrate metabolic process, succinate metabolic process, photosynthesis, cell wall organization, leaf development, triterpenoid biosynthetic process, alpha-amino acid metabolic process, lysine biosynthetic process, monosaccharide metabolic process, indole-containing compound metabolic process (Table S8). Then, we found the terms relevant to cell wall organization, anion transport, external encapsulating structure organization, inorganic anion transport, cell wall organization or biogenesis, ion transport, triterpenoid metabolic process, regulation of localization, regulation of transport, male meiosis, succinate metabolic process, uridine metabolic process, photoperiodism, triterpenoid catabolic process, pentacyclic triterpenoid catabolic process, triterpenoid biosynthetic process, terpenoid catabolic process, terpenoid biosynthetic process and tetracyclic triterpenoid metabolic process were significantly enriched based on DEGs from T2 vs. CK (Table S8). In “Biological Process” between T3 and CK, we found the DEGs in gene expression, cell wall organization, polysaccharide catabolic process, carbohydrate catabolic process, triterpenoid metabolic process, cell wall organization or biogenesis, succinate metabolic process, triterpenoid biosynthetic process, triterpenoid catabolic process, pentacyclic triterpenoid catabolic process, pentacyclic triterpenoid metabolic process, pentacyclic triterpenoid biosynthetic process, inorganic anion transport, external encapsulating structure organization, anion transport, male meiosis and phloem or xylem histogenesis all showed significiant differences. (Table S8; Additional file 5: Table S5). Of note, the number of terms relevant to terpenoid synthesis in T2 vs. CK and T3 vs. CK were higher than that in T1 vs. CK, suggesting that MeJA might mainly caused terpenoid synthesis at 36 or 48 hat in EK.
MeJA altering the transcription pattern of genes involved in numerous pathways represented by terpenoid biosynthesis in Euphorbia kansui.
To investigate the effects of MeJA in affecting the expression of genes involved in biosynthesis in EK, all DEGs were then mapped in the KEGG database (Fig. 4A). For T1 vs. CK comparison, the DEGs mainly involved in Cutin, suberine and wax biosynthesis, Plant hormone signal transduction, Photosynthesis - antenna proteins, alpha-Linolenic acid metabolism, Ribosome, Porphyrin and chlorophyll metabolism, Photosynthesis, Glycosaminoglycan degradation, Phenylalanine, tyrosine and tryptophan biosynthesis, Zeatin biosynthesis, Ubiquinone and other terpenoid-quinone biosynthesis, Cyanoamino acid metabolism, RNA transport, Starch and sucrose metabolism and Phenylalanine metabolism (Fig. 4A; Table S9). Typically, no terms relevant to terpenoid biosynthesis were significantly enriched in T1 vs. CK comparison, suggesting that MeJA treatment did not affect terpenoid biosynthesis at 24 hat in EK (Fig. 4A; Table S9). Then, we found that the DEGs in T2 vs. CK comparison mainly involved in Ribosome, Cutin, suberine and wax biosynthesis, alpha-Linolenic acid metabolism, Fatty acid elongation, Vitamin B6 metabolism, Sesquiterpenoid and triterpenoid biosynthesis, Terpenoid backbone biosynthesis, Diterpenoid biosynthesis and Monoterpenoid biosynthesis (Fig. 4A; Table S9; Additional file 6: Table S6). Numerous terpenoid-related biosynthesis including Sesquiterpenoid and triterpenoid biosynthesis, Terpenoid backbone biosynthesis, Diterpenoid biosynthesis and Monoterpenoid biosynthesis were identified in T2 vs. CK comparison, suggesting that 36 hat of MeJA treatment could triggered the terpenoid biosynthesis in EK. For T3 vs. CK comparison, 12 pathways were significantly enriched, including Ribosome, Fatty acid elongation, Cutin, suberine and wax biosynthesis, Pentose and glucuronate interconversions, Ether lipid metabolism, Vitamin B6 metabolism, Sesquiterpenoid and triterpenoid biosynthesis, Photosynthesis-antenna proteins, Histidine metabolism, Flavonoid biosynthesis, Photosynthesis and Glycerolipid metabolism (Fig. 4A; Table S9). This results about T3 vs. CK also showed that 48 hat of MeJA treatment did not exerted effective activation on terpenoid biosynthesis in EK (Fig. 4A; Table S9). Additionally, KEGG analysis on DEGs from T2 vs. T1 showed that 32 (0.7%) DEGs were identified in “Sesquiterpenoid and triterpenoid biosynthesis”, 37 (0.81%) DEGs were in “Cutin, suberine and wax biosynthesis”, 36 (0.79%) DEGs were in “Terpenoid backbone biosynthesis”, 126 (2.76%) DEGs were in “Plant-pathogen interaction”, 15 (0.4%) DEGs were in “Flavonoid biosynthesis” (0.45%) (Additional file 7: Table S7). Importantly, as shown in Fig. 4B, we found that after 36 hat MeJA treatment, besides Unigene079832, most DEGs encoding key enzymes involved in MVA-like terpenoid biosynthesis were upregulated in EK in comparison with samples from other three experimental groups.
In addition, from KEGG analysis, we found some unigenes in the “Cutin, suberine and wax biosynthesis”, “Flavonoid biosynthesis” pathway expressed differently under MeJA elicitation. Typically, 25 (0.71%), 28 (0.74%), 11 (0.79%), 37 (0.81%), 20 (0.77%) and 28 (0.89%) DEGs respectively in “Cutin, suberine and wax biosynthesis” pathway through pairwise comparison (CK-vs-T1, CK-vs-T2, CK-vs-T3, T1-vs-T2, T1-vs-T3, T2-vs-T3) (Table 2). From Table 3, we found the expression levels of unigenes coding cytochrome P450-like gene including unigene016395, unigene016396, unigene053539 and unigene010561 are all upregulated after MeJA treatment 36 h. Interestingly, we found “Flavonoid biosynthesis” pathway, genes coding chalcone–flavonone isomerase (CHI), shikimate O-hydroxycinnamoyltransferase (HCT), caffeoyl CoA 3-O-methyltransferase (CCOMT) and flavanone 3-hydroxylase (F3H) are all downregulated after MeJA treatment 24 h, however, upregulated at 36 h (Table 3). The expression levels of CHI and F3H reached the highest at 48 h. Taken together, these results suggested that exogenous application of MeJA treatment mainly triggered the terpenoid biosynthesis, especially MVA pathway in EK at 36 hat, while MeJA treatment also altered the transcription pattern of genes involved in biosynthesis of cutin, wax and flavonoid, implying the effects of MeJA in promoting the accumulation of bioactive metabolites in EK.
Table 2
The number of MeJA-responsive DEGs in terpenoid biosynthesis, “Cutin, suberine and wax biosynthesis”, “Flavonoid biosynthesis” pathway of KEGG annotation
| Pathway | DEGs with pathway annotation |
| CK-vs-T1 3539(100%) | CK-vs-T2 3781(100%) | CK-vs-T3 1394(100%) | T1-vs-T2 4568(100%) | T1-vs-T3 2605(100%) | T2-vs-T3 3161(100%) |
| Terpenoid backbone biosynthesis | 40(1.13%) | 45(1.19) | 18(1.29%) | 36(0.79%) | 28 (1.07%) | 33 (1.04%) |
| Ubiquinone and other terpenoid-quinone biosynthesis | 21(0.59%) | 39 (1.03%) | 8 (0.57%) | 39 (0.85%) | 28 (1.07%) | 35 (1.11%) |
| Steroid biosynthesis | 25 (0.71%) | 31 (0.82%) | 5 (0.36%) | 36 (0.79%) | 22 (0.84%) | 32 (1.01%) |
| Sesquiterpenoid and triterpenoid biosynthesis | 14 (0.4%) | 26 (0.69%) | 8 (0.57%) | 32(0.7%) | 16 (0.61%) | 25 (0.79%) |
| Diterpenoid biosynthesis | 8 (0.23%) | 7 (0.19%) | 4 (0.29%) | 11 (0.24%) | 5 (0.19%) | 5 (0.16%) |
| Monoterpenoid biosynthesis | 4 (0.11%) | 3 (0.08%) | 1 (0.07%) | 5 (0.11%) | 3 (0.12%) | 1 (0.03%) |
| Cutin, suberine and wax biosynthesis | 25 (0.71%) | 28 (0.74%) | 11 (0.79%) | 37 (0.81%) | 20 (0.77%) | 28 (0.89%) |
| Flavonoid biosynthesis | 11 (0.31%) | 15 (0.4%) | 14 (1%) | 21 (0.46%) | 13 (0.5%) | 12 (0.36%) |
Table 3
KEGG pathway analysis of the MeJA-responsive differential expressed genes in “Terpenoid biosynthesis”, “Cutin, suberine and wax biosynthesis” and “Flavonoid biosynthesis” pathway
| pathway Enzyme | Unigen ID | Enzyme ID | Pathway ID | fpmk changes of DEG |
| Terpenoid biosynthesis | | | | |
| Terpenoid backbone biosynthesis | | | | |
| MVA | | | | CK 24h 36h 48h |
| acetyl-CoA acetyltransferase | Unigene064228 | EC:2.3.1.9 | map00900 | 0.2 | 0.68 | 2.47 | 0.35 |
| acetyl-CoA acetyltransferase | Unigene022256 | EC:2.3.1.9 | map00900 | 0.2 | 0.68 | 2.47 | 0.35 |
| cytosolic 3-hydroxy-3-methylglutaryl-CoA synthase | Unigene028845 | EC:2.3.3.10 | map00900 | 0.19 | 0.86 | 3.025 | 0.55 |
| 3-hydroxy-3-methylglutaryl-coenzyme A reductase 1-like | Unigene016616 | EC:1.1.1.34 | map00900 | 22.65 | 49.635 | 168.25 | 41.005 |
| probable phosphomevalonate kinase | Unigene078778 | EC:1.1.1.34 | map00900 | 0.001 | 0.085 | 1.08 | 0.09 |
| hydroxymethylglutaryl-CoA reductase | Unigene090341 | EC:1.1.1.34 | map00900 | 0.195 | 0.41 | 1.345 | 0.36 |
| phosphomevalonate kinase isoform X2 | Unigene078776 | EC:2.7.4.2 | map00900 | 0.11 | 0.705 | 1.5 | 0.25 |
| GHMP kinase family protein | Unigene083442 | EC:2.7.4.2 | map00900 | 1.42 | 1.315 | 5.21 | 1.955 |
| phosphomevalonate kinase-like | Unigene083438 | EC:2.7.4.2 | map00900 | 1.895 | 2.575 | 6.52 | 4.105 |
| mevalonate pyrophosphate decarboxylase | Unigene040754 | EC:4.1.1.33 | map00900 | 0.105 | 0.525 | 2.105 | 0.365 |
| isopentenyl diphosphate isomerase | Unigene011459 | EC:5.3.3.2 | map00900 | 0.175 | 0.615 | 2.585 | 0.385 |
| isopentenyl pyrophosphate | Unigene019565 | EC:5.3.3.2 | map00900 | 0.001 | 0.915 | 3.16 | 0.585 |
| geranyl diphosphate synthase | Unigene037391 | EC:2.5.1.1 | map00900 | 0.001 | 0.001 | 0.785 | 0.001 |
| decaprenyl-diphosphate synthase subunit 1 | Unigene003040 | EC:2.5.1.1 | map00900 | 0.001 | 0.14 | 0.465 | 0.14 |
| farnesyl pyrophosphate synthase | Unigene028555 | EC:2.5.1.1 | map00900 | 0.045 | 0.515 | 1.55 | 0.285 |
| farnesyl pyrophosphate synthase 1-like isoform X1 | Unigene079082 | EC:2.5.1.1 | map00900 | 7.955 | 5.275 | 28.985 | 12.605 |
| MEP | | | | | | | |
| probable 1-deoxy-D-xylulose-5-phosphate synthase, chloroplastic isoform X3 | Unigene044979 | EC:2.2.1.7 | map00900 | 0.27 | 0.001 | 1.46 | 0.185 |
| probable 1-deoxy-D-xylulose-5-phosphate synthase 2 | Unigene042075 | EC:2.2.1.7 | map00900 | 28.01 | 11.845 | 6.415 | 4.815 |
| probable 1-deoxy-D-xylulose-5-phosphate synthase 2 | Unigene093405 | EC:2.2.1.7 | map00900 | 70.425 | 26.575 | 20.93 | 10.91 |
| probable 1-deoxy-D-xylulose-5-phosphate synthase 2 | Unigene089680 | EC:2.2.1.7 | map00900 | 76.88 | 25.2 | 25.19 | 11.31 |
| 1-deoxy-D-xylulose-5-phosphate synthase 2 | Unigene089682 | EC:2.2.1.7 | map00900 | 163.78 | 64.69 | 39.105 | 30.01 |
| 4-hydroxy-3-methylbut-2-en-1-yl diphosphate synthase | Unigene056879 | EC:1.17.7.1 | map00900 | 262.015 | 137.31 | 125.12 | 148.96 |
| 4-hydroxy-3-methylbut-2-enyl diphosphate reductase | Unigene003422 | EC:1.17.7.2 | map00900 | 1.915 | 0.3 | 0.001 | 0.69 |
| 4-hydroxy-3-methylbut-2-enyl diphosphate reductase | Unigene003418 | EC:1.17.7.2 | map00900 | 361.155 | 237.77 | 171.695 | 195.195 |
| 2-C-methyl-D-erythritol 4-phosphate cytidylyltransferase | Unigene024815 | EC:2.7.7.60 | map00900 | 1.76 | 0.001 | 0.85 | 0.2 |
| geranylgeranyl pyrophosphate synthase | Unigene016497 | EC:2.5.1.1 | map00900 | 138.865 | 20.76 | 24.02 | 15.325 |
| farnesyl diphosphate synthase | Unigene004541 | EC:2.5.1.1 | map00900 | 25.005 | 12.3 | 11.39 | 9.685 |
| Sesquiterpenoid and triterpenoid biosynthesis | | | | | | | |
| squalene synthase | Unigene032992 | EC:2.5.1.21 | map00909 | 0.08 | 0.39 | 1.47 | 0.19 |
| sucrose synthase | Unigene043983 | EC:2.5.1.21 | map00909 | 31.355 | 19.67 | 40.29 | 16.84 |
| squalene monooxygenase 1, | Unigene014678 | EC:1.14.13.132 | map00909 | 0.17 | 0.895 | 2.525 | 0.38 |
| Squalene monooxygenase | Unigene076528 | EC:1.14.13.132 | map00909 | 2.42 | 10.805 | 2.62 | 3.985 |
| squalene epoxidase | Unigene048572 | EC:1.14.13.132 | map00909 | 3.095 | 7.005 | 3.085 | 2.945 |
| Squalene monooxygenase | Unigene025852 | EC:1.14.13.132 | map00909 | 0.23 | 0.05 | 2.78 | 0.51 |
| squalene monooxygenase-like | Unigene041706 | EC:1.14.13.132 | map00909 | 0.965 | 0.72 | 2.175 | 1.13 |
| beta-amyrin synthase-like | Unigene022072 | EC:5.499.39 | map00909 | 2.575 | 25.455 | 2.73 | 4.57 |
| Steroid biosynthesis | | | | | | | |
| squalene synthase | Unigene032992 | EC:2.5.1.21 | map00100 | 0.08 | 0.39 | 1.47 | 0.19 |
| sucrose synthase | Unigene043983 | EC:2.5.1.21 | map00100 | 31.355 | 19.67 | 40.29 | 16.84 |
| mitogen-activated protein kinase kinase kinase YODA | Unigene043985 | EC:2.5.1.21 | map00100 | 19.57 | 18.49 | 27.07 | 24.29 |
| squalene monooxygenase 1, partial | Unigene014678 | EC:1.14.13.132 | map00100 | 0.17 | 0.895 | 2.525 | 0.38 |
| Squalene monooxygenase | Unigene076528 | EC:1.14.13.132 | map00100 | 2.42 | 10.805 | 2.62 | 3.985 |
| squalene epoxidase | Unigene048572 | EC:1.14.13.132 | map00100 | 3.095 | 7.005 | 3.085 | 2.945 |
| N(6)-adenine-specific DNA methyltransferase 2 | Unigene072562 | EC:1.14.13.132 | map00100 | 0.12 | 0.395 | 1.07 | 0.265 |
| N(6)-adenine-specific DNA methyltransferase 2 | Unigene002085 | EC:1.1.1.170 | map00100 | 55.41 | 251.2 | 42.07 | 82.475 |
| delta(24)-sterol reductase-like | Unigene014530 | EC:1.3.1.72 | map00100 | 0.155 | 0.845 | 2.135 | 0.375 |
| 3beta-hydroxysteroid-dehydrogenase/decarboxylase isoform | Unigene022670 | EC:1.1.1.170 | map00100 | 0.185 | 0.9 | 2.615 | 0.29 |
| cycloartenol-C-24-methyltransferase 1-like | Unigene036530 | EC:ERG6 | map00100 | 0.185 | 1.04 | 2.53 | 0.33 |
| 7-dehydrocholesterol reductase | Unigene020078 | EC:1.3.1.21 | map00100 | 0.565 | 2.4 | 8.355 | 1.245 |
| acyl-coenzyme A:cholesterol acyltransferase | Unigene013755 | EC:2.3.1.26 | map00100 | 0.07 | 0.355 | 0.98 | 0.305 |
| cycloartenol synthase | Unigene050717 | EC:5.4.99.8 | map00100 | 2.63 | 7.885 | 2.565 | 1.845 |
| cycloartenol-C-24-methyltransferase 1-like | Unigene036530 | EC:SMT1 | map00100 | 0.185 | 1.04 | 2.53 | 0.33 |
| Obtusifoliol 14alpha-Demethylase | Unigene061050 | EC:CYP51G1 | map00100 | 0.075 | 0.615 | 2.245 | 0.265 |
| C-4 sterol methyl oxidase (ISS) | Unigene082000 | EC:SMO2 | map00100 | 0.11 | 0.875 | 3.055 | 0.485 |
| C-5 sterol desaturase | Unigene002420 | EC:1.14.21.6 | map00100 | 0.195 | 0.445 | 1.795 | 0.265 |
| triacylglycerol lipase | Unigene081843 | EC:3.1.1.13 | map00100 | 0.001 | 0.915 | 2.215 | 0.001 |
| triacylglycerol lipase | Unigene031929 | EC:3.1.1.13 | map00100 | 0.13 | 0.42 | 1.045 | 0.16 |
| esterase / lipase | Unigene009853 | EC:3.1.1.13 | map00100 | 0.12 | 0.12 | 0.91 | 0.1 |
| Monoterpenoid biosynthesis | | | | | | | |
| F-box/kelch-repeat protein At3g23880-like | Unigene086409 | EC:1.1.1.206 | map00902 | 0.001 | 1.265 | 0.001 | 0.001 |
| probable terpene synthase 11 | Unigene055596 | EC:4.2.3.25 | map00902 | 0.965 | 0.001 | 0.13 | 0.001 |
| (+)-neomenthol dehydrogenase-like isoform X1 | Unigene019800 | EC:1.1.1.208 | map00902 | 1.89 | 0.19 | 2.155 | 2.265 |
| (+)-neomenthol dehydrogenase-like | Unigene092575 | EC:1.1.1.208 | map00902 | 2.32 | 0.26 | 6.075 | 1.835 |
| Diterpenoid biosynthesis | | | | | | | |
| ent-kaurene oxidase | Unigene014957 | EC:1.14.13.78 | map00904 | 0.11 | 0.33 | 1.13 | 0.205 |
| cytochrome P450 82G1-like | Unigene019714 | EC:CYP82G1 | map00904 | 0.72 | 0.145 | 2.66 | 0.68 |
| ent-kaur-16-ene synthase | Unigene023377 | EC:4.2.3.19 | map00904 | 47.29 | 2.56 | 1.875 | 3.81 |
| gibberellin 3-beta-dioxygenase 1 | Unigene040056 | EC:1.14.11.15 | map00904 | 0.14 | 0.14 | 1.725 | 0.425 |
| gibberellin 2-beta-dioxygenase-like | Unigene068133 | EC:1.14.11.13 | map00904 | 32.09 | 48.075 | 15.57 | 31.55 |
| ent-copalyl diphosphate synthase | Unigene083812 | EC:5.5.1.13 | map00904 | 3.34 | 0.16 | 0.245 | 0.195 |
| ent-kaurene synthase | Unigene091248 | EC:4.2.3.19 | map00904 | 4.795 | 0 | 2.17 | 1.82 |
| Ubiquinone and other terpenoid-quinone biosynthesis | | | | | | | |
| homogentisate geranylgeranyl transferase | Unigene042099 | EC:25.1.115 | map00130 | 1.11 | 3.485 | 0.38 | 1.08 |
| homogentisate solanesyltransferase, chloroplastic | Unigene080040 | EC:25.1.117 | map00130 | 5.025 | 4.71 | 12.035 | 10.125 |
| 37 kDa chloroplast inner membrane protein | Unigene059865 | EC:2.1.1.295 | map00130 | 1 | 0.915 | 11.99 | 2.625 |
| 37 kDa inner envelope membrane protein | Unigene039484 | EC:2.1.1.295 | map00130 | 2.32 | 0.72 | 10.325 | 4.495 |
| 2-methyl-6-phytyl-1,4-hydroquinone methyltransferase, chloroplastic | Unigene077544 | EC:2.1.1.295 | map00130 | 14.835 | 3.505 | 44.745 | 22.845 |
| gamma-tocopherol methyltransferase | Unigene025639 | EC:2.1.1.95 | map00130 | 8.485 | 5.305 | 17.89 | 10.695 |
| probable tocopherol O-methyltransferase, chloroplastic isoform X2 | Unigene069994 | EC:2.1.1.95 | map00130 | 18.325 | 18.54 | 36.685 | 26.95 |
| 4-hydroxybenzoate polyprenyl transferase | Unigene067103 | EC:Coq2 | map00130 | 0.07 | 0.375 | 0.71 | 0.12 |
| 2-methoxy-6-polyprenyl-1,4-benzoquinol methylase, mitochondrial | Unigene080019 | EC:Coq5 | map00130 | 0.001 | 2.675 | 0.275 | 0.001 |
| 2-methoxy-6-polyprenyl-1,4-benzoquinol methylase, mitochondrial | Unigene056630 | EC:Coq5 | map00130 | 0.13 | 0.465 | 1.525 | 0.14 |
| ubiquinone biosynthesis protein COQ7 | Unigene084930 | EC:Coq7 | map00130 | 0.08 | 0.575 | 1.91 | 0.375 |
| retrovirus-related Pol polyprotein from transposon TNT 1–94 | Unigene063000 | EC:6.2.1.26 | map00130 | 0.77 | 2.17 | 0.15 | 0.62 |
| 1,4-dihydroxy-2-naphthoyl-CoA synthase, peroxisomal-like | Unigene070519 | EC:4.1.3.36 | map00130 | 10.16 | 26.055 | 11.115 | 10.055 |
| Hexaprenyldihydroxybenzoate methyltransferase | Unigene013837 | EC:2.1.1.114 | map00130 | 0.18 | 0.63 | 1.2 | 0.001 |
| protein PHYLLO, chloroplastic isoform X3 | Unigene024447 | EC:5.4.4.2 | map00130 | 0.245 | 0.62 | 2.8 | 0.775 |
| protein PHYLLO, chloroplastic | Unigene095857 | EC:5.4.4.2 | map00130 | 0.82 | 0.52 | 3.74 | 1.305 |
| protein PHYLLO, chloroplastic | Unigene064679 | EC:5.4.4.2 | map00130 | 0.72 | 0.41 | 1.98 | 0.74 |
| isochorismate synthase 2, chloroplastic-like isoform X1 | Unigene012041 | EC:5.4.4.2 | map00130 | 29.525 | 22.26 | 105.14 | 51.745 |
| isochorismate synthase 2, chloroplastic | Unigene012036 | EC:5.4.4.2 | map00130 | 20.345 | 9.515 | 45.455 | 29.97 |
| isochorismate synthase 2, chloroplastic | Unigene012037 | EC:5.4.4.2 | map00130 | 24.01 | 13.685 | 51.935 | 40.1 |
| geranylgeranyl diphosphate reductase | Unigene043490 | EC:1.3.1.83 | map00900 | 46.05 | 27.115 | 101.745 | 92.595 |
| Cutin, suberine and wax biosynthesis | | | | | | | |
| cytochrome P450 86A8-like | Unigene061395 | EC:CYP86A4 | map00073 | 0.63 | 2.755 | 10.23 | 1.745 |
| cytochrome P450 86A8-like | Unigene061396 | EC:CYP86A4 | map00073 | 1.935 | 5.065 | 24.21 | 2.73 |
| Os04g0560100 | Unigene010561 | EC:CYP86A4 | map00073 | 0.14 | 0.001 | 1.355 | 0.06 |
| cytochrome P450 86A7 | Unigene053539 | EC:CYP86A4 | map00073 | 1.465 | 3.84 | 10.82 | 1.67 |
| Flavonoid biosynthesis | | | | | | | |
| chalcone–flavonone isomerase | Unigene040233 | EC:5.5.1.6 | map00941 | 19.97 | 7.475 | 26.43 | 27.19 |
| shikimate O-hydroxycinnamoyltransferase | Unigene048505 | EC:2.3.1.133 | map00941 | 2.825 | 0.415 | 22.77 | 6.655 |
| shikimate O-hydroxycinnamoyltransferase | Unigene022901 | EC:2.3.1.133 | map00941 | 4.48 | 0.665 | 13.565 | 8.785 |
| shikimate O-hydroxycinnamoyltransferase | Unigene070081 | EC:2.3.1.133 | map00941 | 1.43 | 0.115 | 1.73 | 3.33 |
| caffeoyl CoA 3-O-methyltransferase | Unigene071049 | EC:2.3.1.74 | map00941 | 1.525 | 0.001 | 1.78 | 0.975 |
| flavanone 3-hydroxylase | Unigene054810 | EC:1.1.4.11.9 | map00941 | 26.505 | 20.38 | 49.09 | 61.335 |
MeJA treatment activated terpenoid biosynthesis and promoted the accumulation of terpenoid in Euphorbia kansui.
In order to experimentally verify the DEG profile obtained by sequence data in terpenoid biosynthesis associated with MeJA treatment, qRT-PCR analysis was conducted on genes relevant to terpenoid biosynthesis in EK following MeJA treatment. A total of 18 genes coding key enzymes in terpenoid biosynthesis pathway, including AACT, HMGS, HMGR, PMK, GHMP, MDC, DXS, MCS, HDS, HDR, IDI, DPPI, FPPS, GPPS, GGPPS, GGPPR, SUS and N6AMT2 (Table 3) were selected to analyze the gene expression pattern under different MeJA treatment (Fig. 5). The gene and β-ACTIN-specific primers for qRT-PCR were listed in Additional file 1: Table S1. The results suggested that the expression levels of the majority of the genes verified were consistent with those from RNA transcriptome data (Fig. 5). Most of these genes relevant to terpenoid biosynthesis were upregulated in EK by MeJA treatment at 36 hat (Fig. 5). Overall, qRT-PCR verified the effects of MeJA treatment in activating the transcription of terpenoid biosynthesis in EK at 36 hat.
Subsequently, we determined the content of the leaf total terpenoid at different MeJA treatment by ultraviolet-visible spectrophotometry. The euphadienol served as standard substance. The results showed that the content of total terpenoid was higher at 24 h 20 µM MeJA treatment to CK, and reached the highest at 36 h µM MeJA treatment, and with the time of MeJA treatment prolonged to 48 h, the content of the total terpenoids decreased (Fig. 6). Compared with the control, the content of total terpenoid was on average about 2.38 times at 24 h MeJA treatment and 2.80 times at 36 h MeJA treatment (Fig. 6). The statistical analysis at MeJA treatments 36 h showed significant differences (P < 0.05) to the control (Fig. 6). Thus, these results confirmed that exogenous application of MeJA could activate terpenoid biosynthesis and promote the accumulation of terpenoid in EK at 36 hat, then leading to high quality of EK for usage.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}