Despite the plethora of reports to clarify the nature of GH19 chitinases from a panel of organisms including plants, viruses, nematodes, etc., still more research is mandatory to elucidate the main structural differences between the unexplored bacterial GH19 chitinases and the well-studied GH19 chitinases of non-bacterial members. Recently, the huge number of microbial genome projects does introduce a hill of complete microbial genome sequences in the international repository nucleotide databases (e.g., GenBank, EMBL, and DDJA) with putative and annotated GH19 chitinases from numerous bacteria. Consequently, the above mentioned does address the indispensable need for the heterologous expression of the putative bacterial GH19 chitinases sequences to better understand the nature and the mode of action of these enzymes.
In our study, the multispecies class I GH19 chitinase namely chiRAM was cloned and expressed in E. coli BL21 (DE3) Rosetta for the first time ever. Blastp analysis conferred a high sequence identitiy ranged from 99.34–91.84 % among protein sequences of GH19 chitinase of different Enterobacter spp. and chiRAM (Table 1). This would in turn explore the rationale behind the nomenclature of the multispecies GH19 chitinase from Enterobacter spp. in Genbank. Likewise GH18 chitinases, the number and configuration of domains of plant GH19 chitinases differ widely in the five classes I, II, IV, VI, and VII [8]. In spite of this discrepancy, each domain has its own unique function that does support the overall chitinase activity or not. The domains of GH19 chitinases include the N-terminal signal peptide region, catalytic domain, chitin binding domain, and C-terminal extension region inferred from the literature of review [9]. A typical CBD in chiRAM belonged to chtBD3 superfamily was evidenced. The chtBD3 superfamily does encompass modules of 40–60 amino acid residues with six conserved aromatic amino acids residues and three hydrophobic amino acids side chains, that help bind chitin and /or cellulose. The present data are in a partial agreement with that of ChBD chiA1 from Bacillus circulans WL-12; with three hydrophobic amino acid residues localized at Val668, Cys677, and Leu695 [39] and ChBDCvChi45 of Chromobacterium violaceium with three hydrophobic amino acid residues at Val41, Ala50, and Leu70 [40]. Moreover, the presence and the position of the CBD region vary widely among different classes (I, II, IV, VI, and VII) of GH 19 chitinases members concluded from the CDD database. The CBD at the N-terminus region is considered one signature feature of class I GH 19 chitinases from plants [41]. In accordance, the absence of expression from the pET-28a(+)/chiRAM-CatDom greatly verified that chiRAM is belonging to class I GH19 chitinases, well known with their CBD as an essential domain for the overall enzyme activity and enzyme structure.
Conversely, members of class I GH19 chitinases from bacterial origin showed a wide divergence in the position of the CBD evidenced from the analysis of the collected sequences from CAZY database. Likewise C-terminus CBD of chiRAM, the C-terminus ChtBD(s) were annotated in the following class I GH19 chitinase sequences: BAE86996.1 chitinase (ChiC_BD: 437–474) [V. proteolyticus], SPM16686.1 putative chitinase (ChiC_BD: 439–479) [V.cholerae], and ACU62980.1 Chitinase (ChtBD3 superfamily: 488–528 & ChiC_BD: 554–595) [Chitinophaga pinensis DSM 2588]. Unlike C-terminus CBD of chiRAM, the N-terminus ChtBD(s) were annotated in the following putative class I GH19 chitinase sequences: APJ14276.1 chitinase ChiC_BD: 19–53 [Aeromonas hydrophila], ACH66624.1 basic endochitinase (ChiC_BD: 24–63) [Aliivibrio fischeri MJ11], ARU51128.1 chitinase (ChiC_BD: 58–100) [Cellulosimicrobium cellulans], AVG16867.1 chitinase (ChiC_BD: 20–59) [Chromobacterium vaccinii], ALN86380.1 secreted chitinase domain protein (ChtBD3 superfamily: 66–113) [Lysobacter capsici], SCF20675.1 chitinase (ChiC_BD: 97–138 & ChtBD3 superfamily: 42–76) [Micromonospora chokoriensis], AAQ24634.1 chitinase (ChiC_BD: 35–74) [Streptomyces griseobrunneus], 1WVU_A chitinase (ChtBD3 superfamily: 6–45) [Str. griseus], ABZ86730.1 chitinase (ChiC_BD: 153–193) [Francisella philomiragia subsp. philomiragia ATCC 25017], and ATP28490.1 chitinase (ChiC_BD: 21–59) [Chromobacterium violaceum]. The deviating sequences and assemblies among diverse groups suggest that they may have evolved from unlike ancestral genes.
Class I plant GH19 chitinases have signature motifs like SHETTG and NYNY. The signature motif SHETTG was found in many three dimensional structural models of plant chitinases (e.g., PDB 3cql endochitinase Carica papaya, PDB 2baa endochitinase Hordeum vulgare, PDB 3iwr chitinase Oryza sativa subsp. Japonica, PDB 4j01 basic endochitinase Secale cereale, PDB 2z37 chitinase Brassica juncea, PDB 4MST chain A, class I chitinase Hevea brasiliensis, and P24091.1 endochitinase B Precursor Nicotiana tabacum). Upon tracing the SHETTG motif in class I GH19 chitinases among bacteria, a modification was noticed in this consensus sequence (Fig. 2B). For instance, the signature motif SHETGG was found in the following sequences: BAA92252.1 chitinase class I Burkholderia gladioli, BAA88833.1 chi35 Str. thermoviolaceus, PDB1WVU Chain A chitinase C Str. griseus, QAT82978.1 class I chitinase Corallococcus coralloides, ABL92611.1 chitinase Mycobacterium sp. KMS and QCW26020.1 chitinase Lysobacter enzymogenes. However, the signature motif AQETGG was observed in the next sequences: APJ14276.1 chitinase A. hydrophila, AWE26318.1 chitinase Salmonella enterica, and CCG31634.1 chitinase Klebsiella aerogenes EA1509E. While the signature motifs AQETGH, SQETTG, and AQETGG were found in BAD92016.1 chitinase I B. circulans, ACU62980.1 chitinase Chitinophaga pinensis DSM 2588, and QCC87378.1 chiRAM chitinase, respectively. In contrast, the NYNY signature motif was fully conserved among class I chitinases from plant and bacteria (Fig. 2B).
The CBD of chiRAM is linked to the catalytic domain by a hing region of 56 residues; rich in proline (Fig .1B). Simialrly, bacterial glycoside hydrolases (GH) like celulases, xylanases and chitinases are chacarerized by the presence of proline or threonine rich spacers connecting the catalytic domain to the non –catalytic binding domain[42]. These spacers or linkers were previously proved to fine tune the binding and the catalytic effieiciny of these enzymes to their insoluble substrates [43]. The modular structure of chiRAM revealed the presence of the unique PKD domain that is completely absent in plant GH18/GH19 chitinases and bacterial GH18 chitinases. Conversely, the PKD domain was annotated in the putative GH19 chitinase sequences of bacteria like (AWE30884.1) of Salmonella enterica, (AXY29321.1) of Klebsiella aerogenes, (ALB71769.1) of Cronobacter muytjensii ATCC 51329, and (ABK38513.1) of Aeromonas hydrophila subsp. hydrophila ATCC 7966. The PKD refers to a protein that is associated with the polycystic kidney syndrome. The function of the PKD domain in these putative GH19 chitinase sequences is still unraveled as these chitinases have not been studied yet. However, its evolution in Enterobacter spp. GH19 chitinases might be attributed to the colonization of Enterobacter in the intestine of human and animals.
Like all secreted proteins, chiRAM has a signal peptide that possibly does initiate the transport across the cytoplasmic membrane. Moreover, the fine structure of chiRAM signal peptide was in a great accordance with those of GH19 chitinases from Aeromonas sp., Str. griseus, and GH18 chitinase of Str. lividance [19]. The signal peptide in the wild type chitinase-producing bacteria plays an essential role to transfer the expressed chitinases from inside the cell to outside the cells as the chitin polymer is too big to be imported into the cell.
Regarding to the structural configuration of chiRAM, the predicted secondary structure and the predicted three dimensional structure assumed its richness in α-helix chains (10 chains). This is quite compatible with the structural folding of plant GH19 chitinases [44] and bacterial GH19 chitinases (e.g., GH19 Pseudomonas aeruginosa) [45] with a conserved core region consisting of 10 α-helices. Conversely, the configuration of GH18 chitinases is in α/β barrel [46].
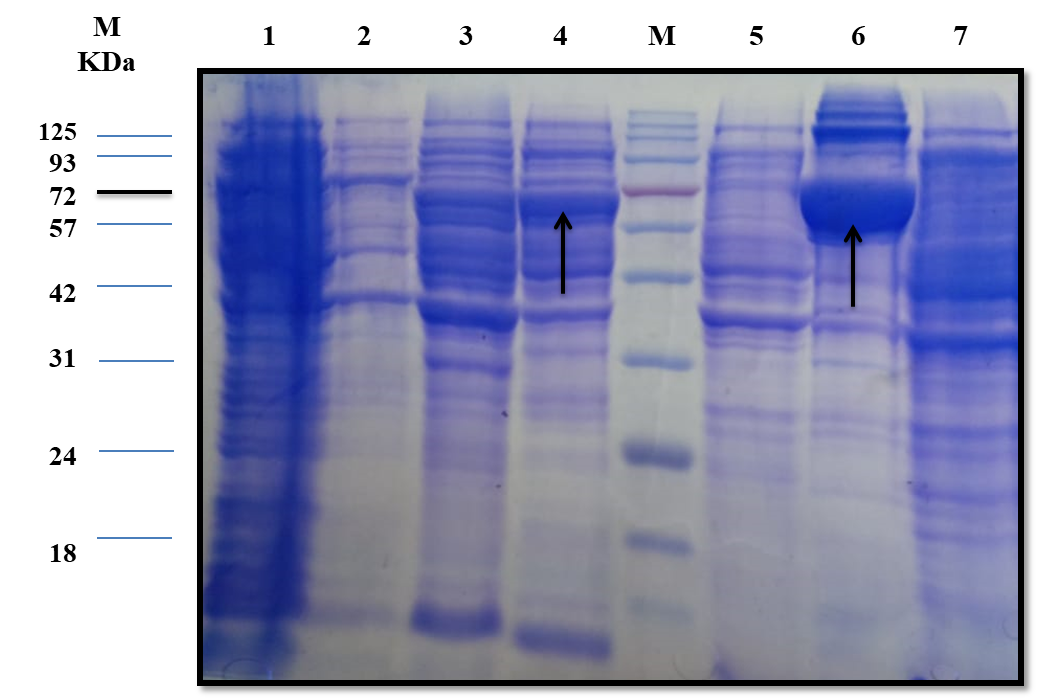
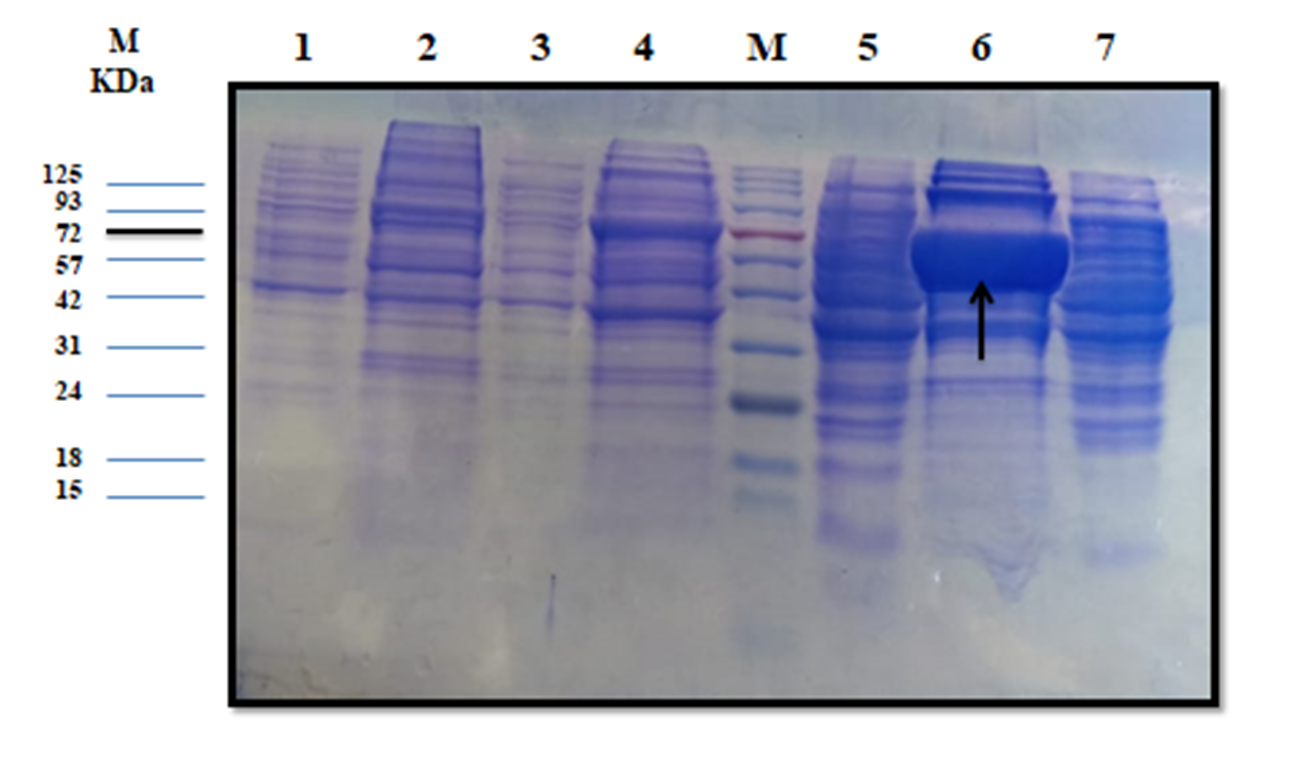
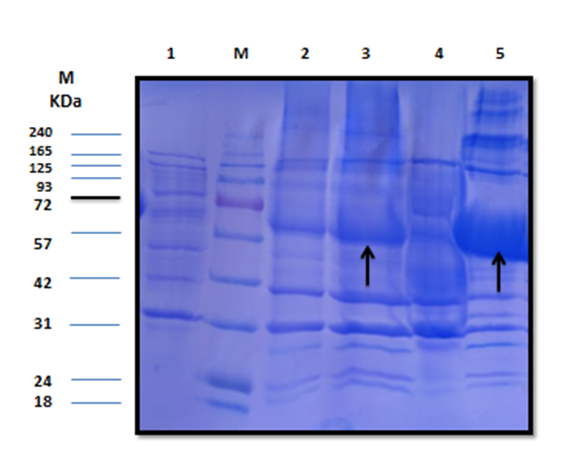
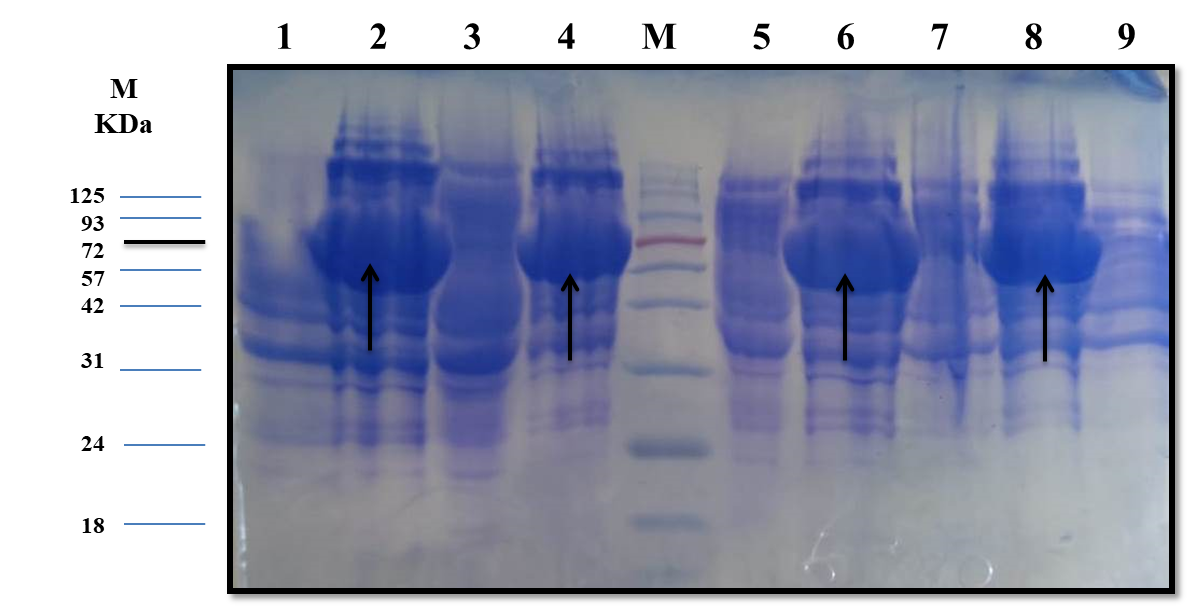
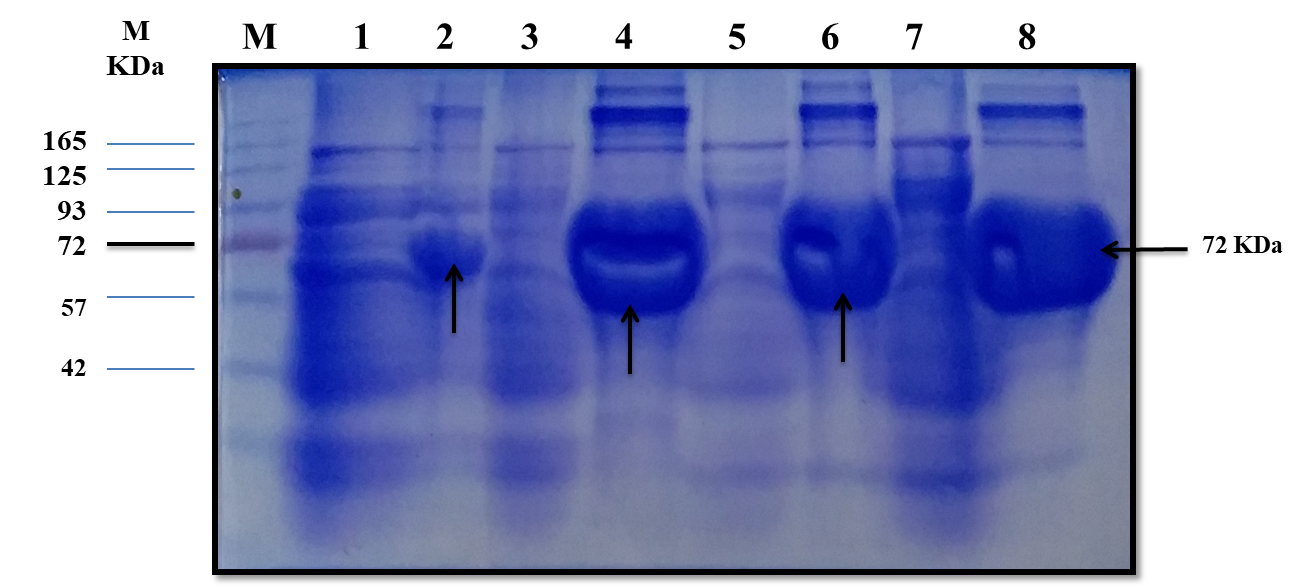
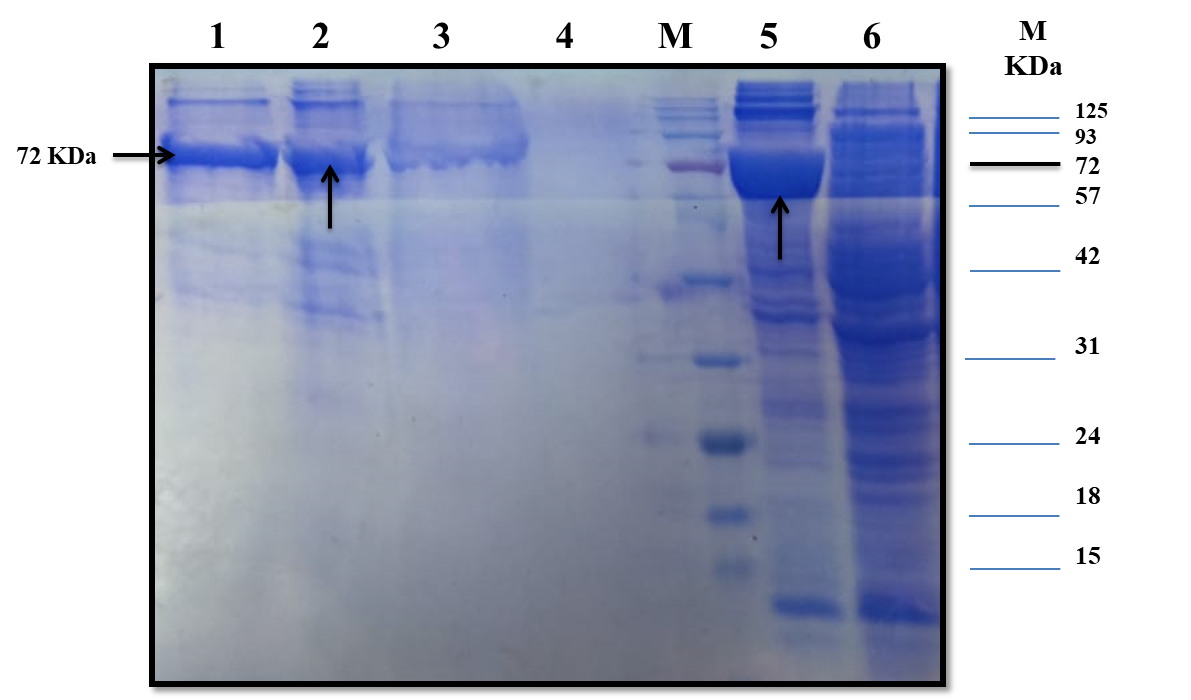
The estimated molecular weight of the recombinant chiRAM (~ 72 kDa) was slightly higher than than the theortical molecualr weight (66 kDa). The increase in the molecular weight of the recombinant chiRAM could be imposed by the presence of his-tags (~ 2.5kDa) [47] and the slow migration of chiRAM on SDS-PAGE with relation to the protein structure of GH19 chitinases [19].
Heterologous expression in the microbial cell factory E.coli is one of the well-established strategies for both small and large scale production of recombinant proteins because E.coli is easy to grow and the overall process is cost-effective. Notwithstanding, the formation of large, insoluble, mis-folded versions of the recombinant proteins in the cytoplasm namely inclusion bodies (IBs) is the main obstacle that hinders the consecution of the overall process, obtaining high levels of active soluble proteins. The exact mechanism controlling the formation of IBs is still not completely understood [48]. However, altered cellular homeostasis would trigger the formation of IBs [48]. The literature of review reported a number of strategies aimed at obtaining soluble, properly folded, active protein species like tailoring cultural conditions (temperature, inducer, and growth rate), genetic manipulation of the target (protein truncation and fusion to solubilization tags), specific codon usage optimization, and exposing the IBs to a panel of solubilization buffers [49]. Nevertheless, there is no a generic method fitting for the solubilization and re-folding of all proteins so far. For each case, there must be a strategy tailored based on trials and errors; custom–made strategies [50]. The initial level of recombinant chiRAM expressed in E.coli BL21 (DE3) Rosetta strain was 0.300 ± 0.0003 U/mL. The applied optimized strategy did successfully enhance the traced chiRAM activity to 1.852 ± 0.0002 and 1.974 ± 0.0002 U/mL in the cell lysate of the recombianant Rosetta strain upon substituting the LB growth medium with M9 and SB, respectively. Herein, a fold enhancement of 6.17 and 6.58 in chiRAM activity expressed by from the recombinant Rosetta strain was obtained upon applying M9 and SB as growth media compared to the yield obtained upon using the conventional LB medium. The compositions of the different media (LB, TB, M9, 2xTY, 5xLB, and SB) do likely trigger an influence on the cellular physiology of the recombinant E.coli (DE3) cells. Consequently, the differences in cellular physiology among different E.coli (DE3) growing on different media most probably impose influences on the nature and the folding status of the expressed recombinant enzymes from these strains. Interestingly, a further enhancement in chiRAM activity from the recombinant Rosetta strain would likely be achieved upon scaling up the process in a laboratory scale fermenter in prospective studies. Conversely, the four solubilzation buffers (B,C, E, and F) could successfully solubilize the chiRAM IBs without tracing any acivity. This could be attributed to the irreversible enzyme denaturation triggered by the denaturants Guanidine-HCl and urea included in these buffers.
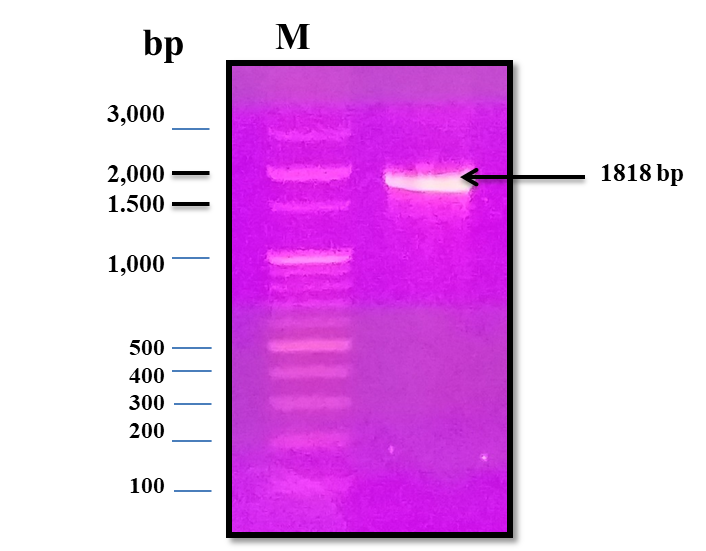
Since a small amount of the recombinant chiRAM was expreesed in the soluble fraction, it was mandatory to get further confirmation about the identity of the band of ~ 72 kDa in the soluble fraction by mass spectrometry. LC-MS-MS confirmed the identity of this band in the soluble fraction as chitinase similar to that of Enterobacter cloacae chitinase/GH19 (WP_063869339.1 & 66.07 kDa) with 52% sequence coverage.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}