The status of AMR in clinic E. coli and S. aureus isolates.
During the sampling period from 2018 to 2022, 2625 clinical isolates of E. coli and 994 clinical isolates of S. aureus were collected. The annual sampling quantity of the two bacteria species was shown in supplement Table 2.
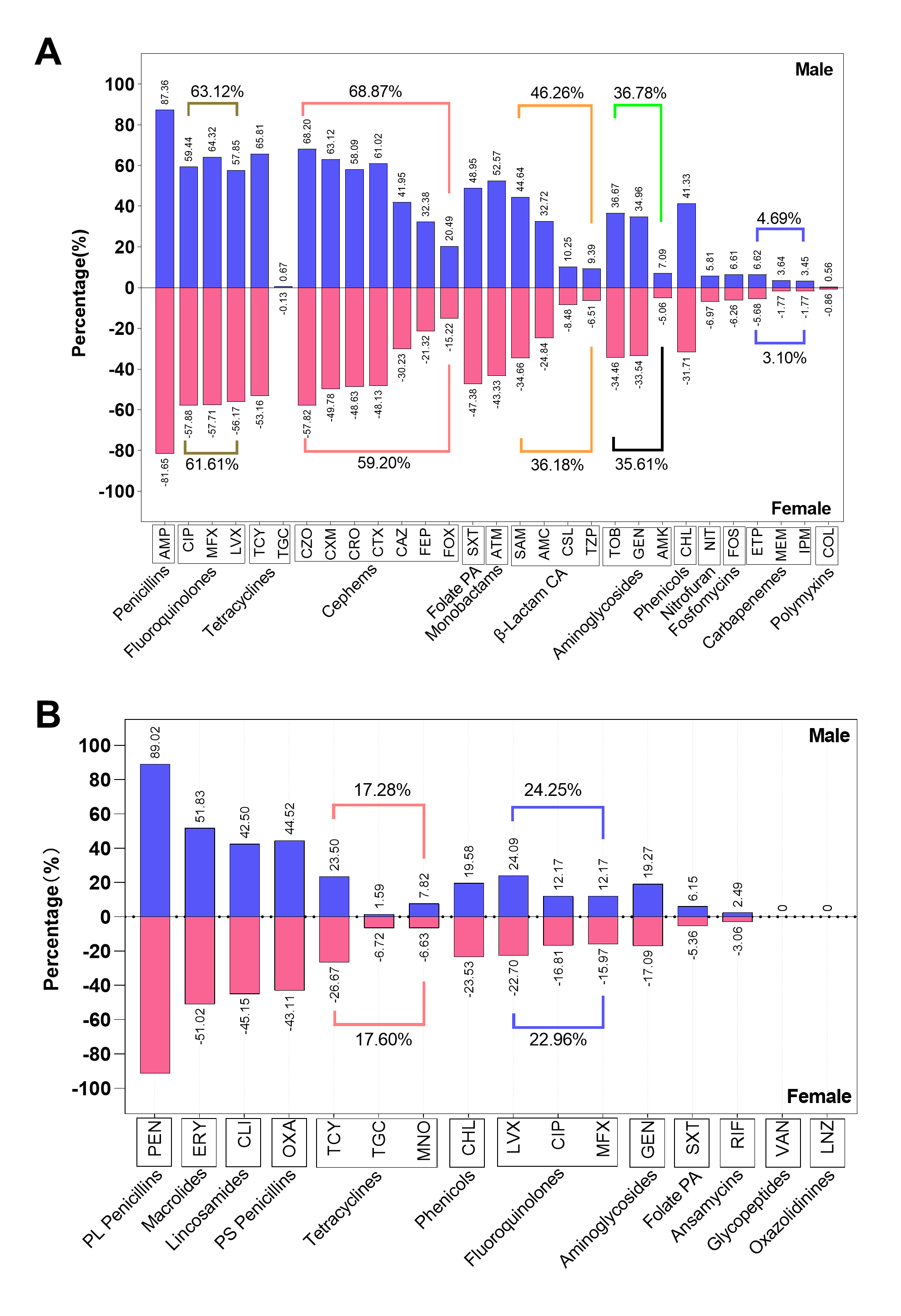
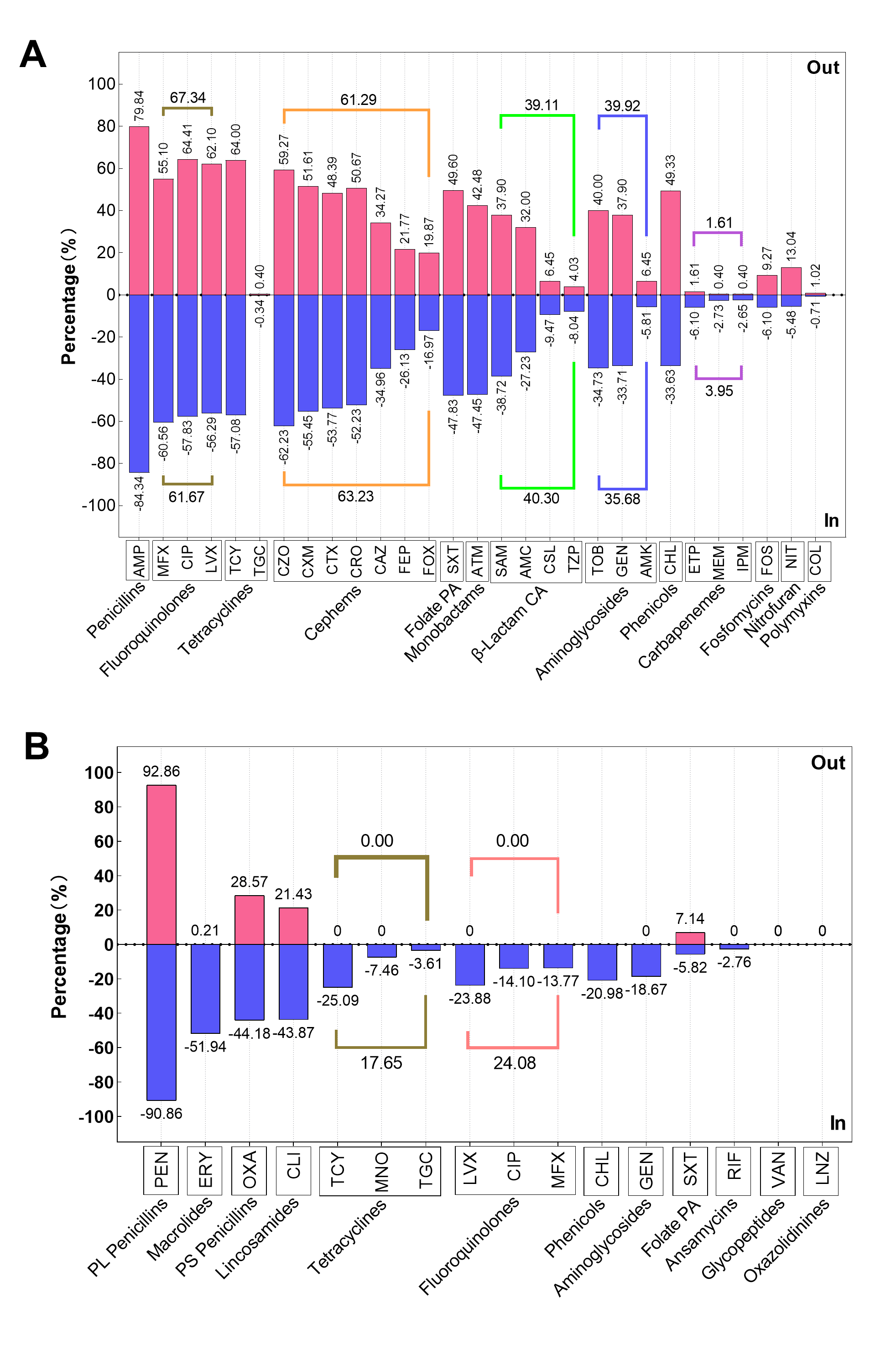
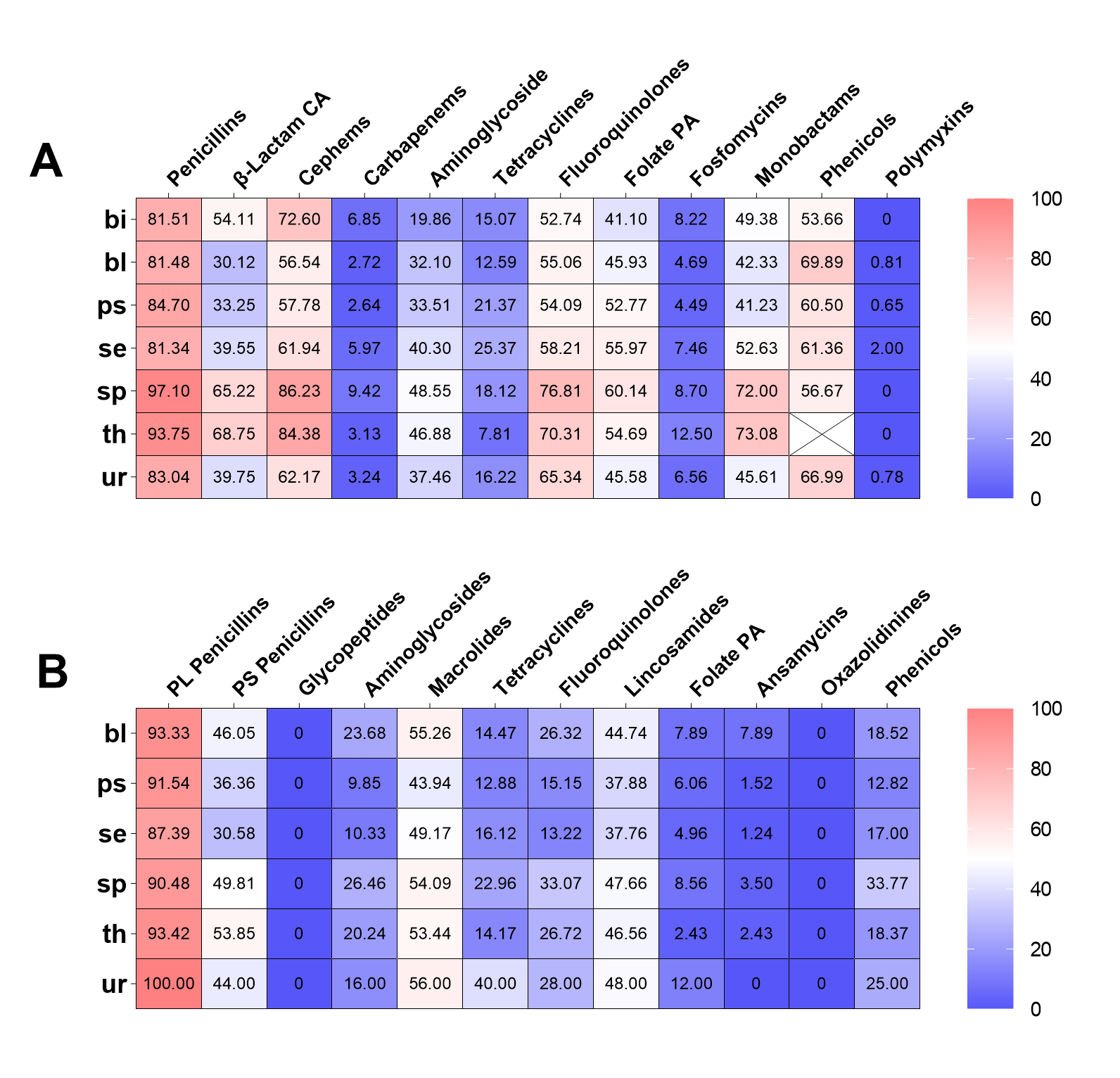
The AST results revealed that a significant proportion of clinical E. coli isolates demonstrated resistance to penicillins (83.92%), cephems (63.05%), fluoroquinolones (62.21%), and tetracyclines (57.77%, Fig. 1A). A low proportion of clinical E. coli isolates were resistant to carbapenems (3.73%), fosfomycin (6.40%), nitrofuran (6.73%), polymyxins (0.74%, Fig. 1A). Out of the six antimicrobial classes that contain more than one agent, the resistance prevalence of distinct agents within the same class was evident discrepant except for classes of carbapenems and fluoroquinolones (Fig. 1C). Among the tetracyclines class, tigecycline exhibited a resistance prevalence nearly zero (0.34%), whereas tetracycline displayed a resistance prevalence exceeding fifty percent (57.77%).Moreover, of the 7 agents classified as cephems, the isolates exhibited the most significant resistance prevalence to first-generation agent (cefazolin [61.95%]), followed by second-generation agents (excluding cefoxitin, cefuroxime [55.09%]) and third-generation agents (cefotaxime [53.26%], ceftazidime [34.90%], and ceftriaxone [52.07%]). The isolates exhibited the lowest resistance prevalence to fourth-generation agents (cefepime [25.71%]).
The clinical S. aureus isolates showed high resistance to penicillinase-labile penicillins (90.89%), macrolides (51.51%), penicillinase-stable penicillins (43.96%), lincosamides (43.55%). Notably, all isolates of S. aureus were susceptible to glycopeptides and oxazolidinines (Fig. 1B). In addition, the clinical S. aureus exhibited low resistance to the folate pathway antagonists (5.84%) and ansamycins (2.72%). All classes of tetracyclines and fluoroquinolones contain three agents in the clinical S. aureus AST, and the resistance prevalence of distinct agents was also discrepant except ciprofloxacin and moxifloxacin (Fig. 1D). Furthermore, tigecycline (3.57%) and minocycline (7.35%), both from the tetracycline class, have resistance prevalence below 10%.
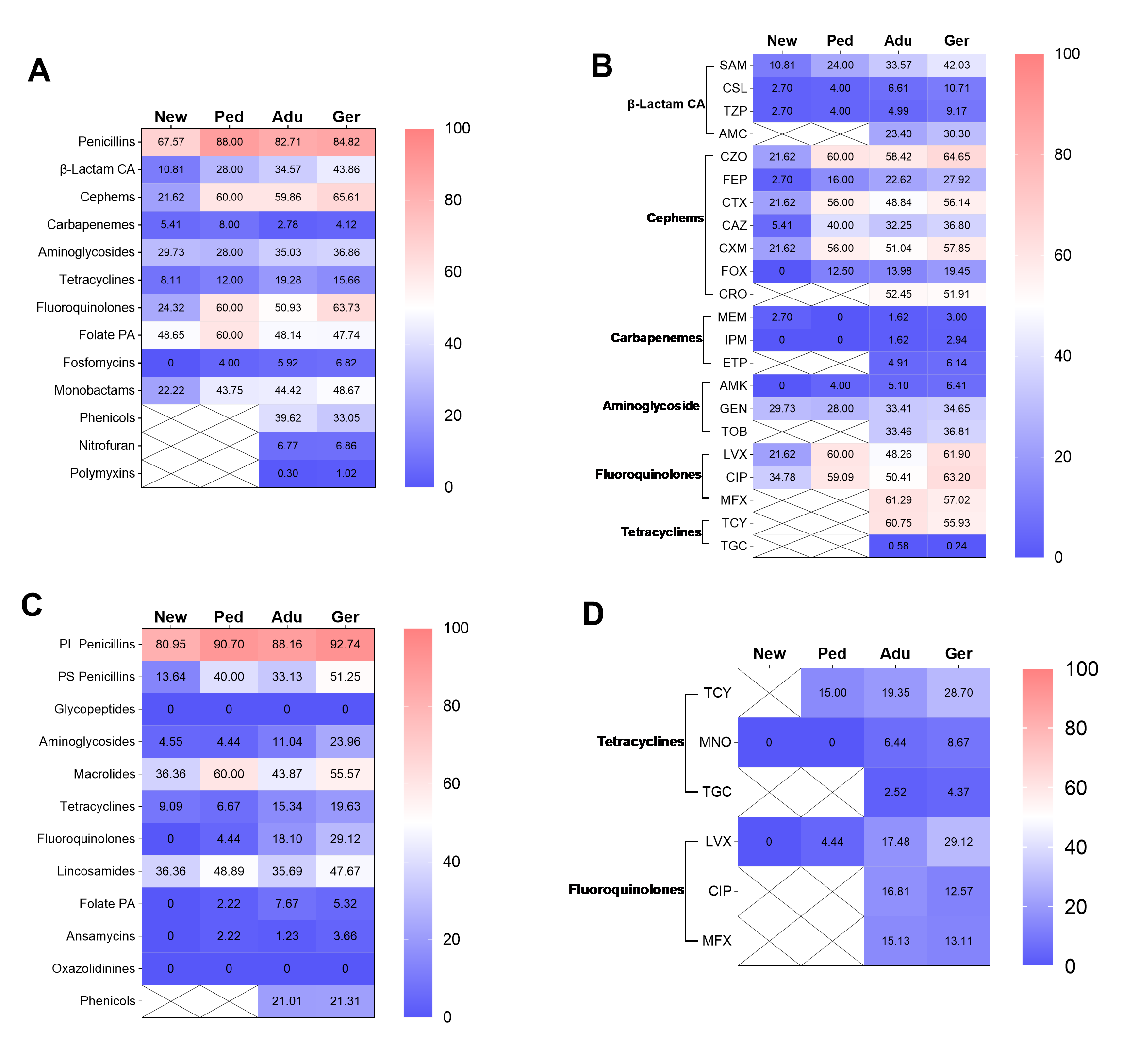
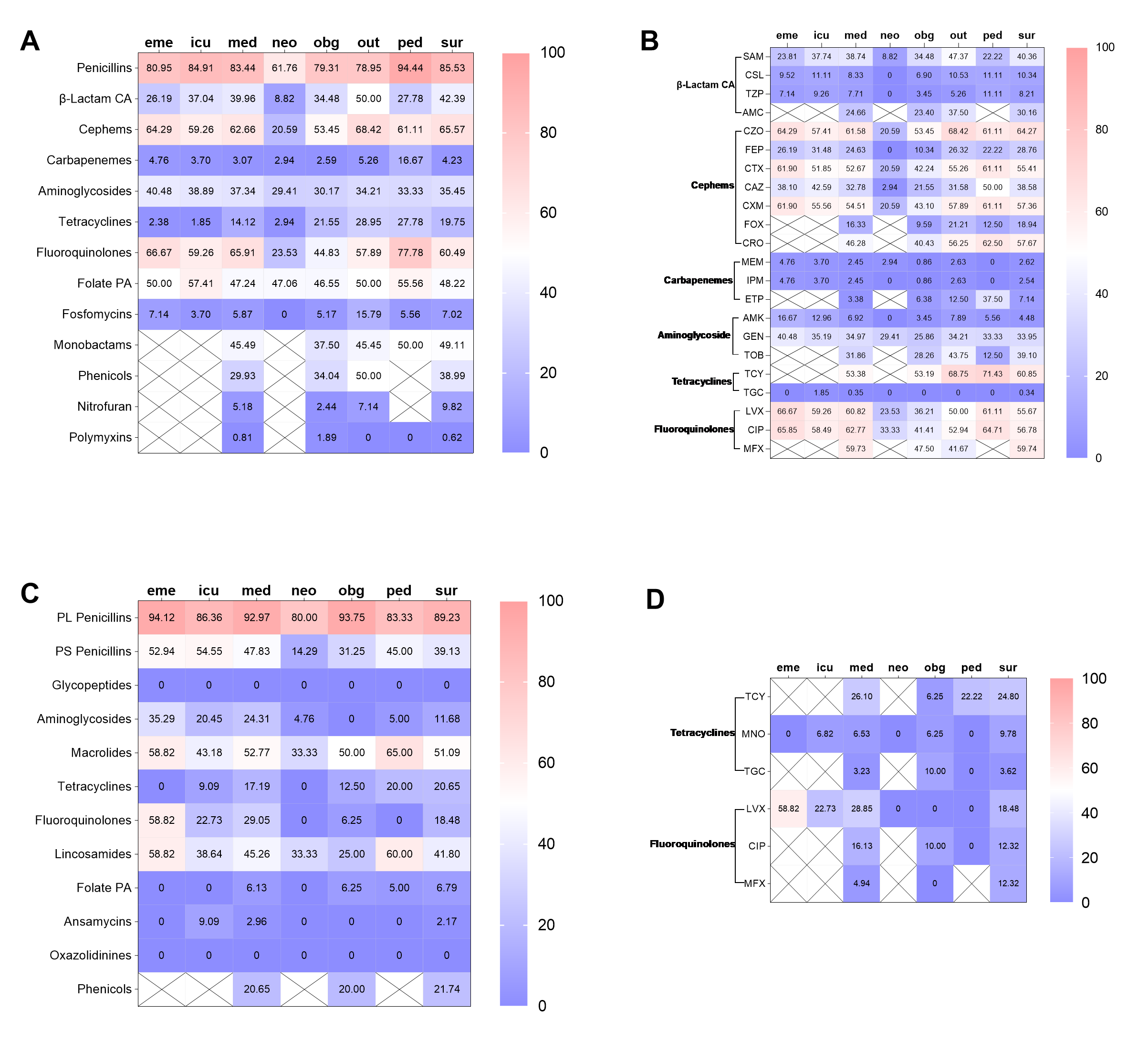
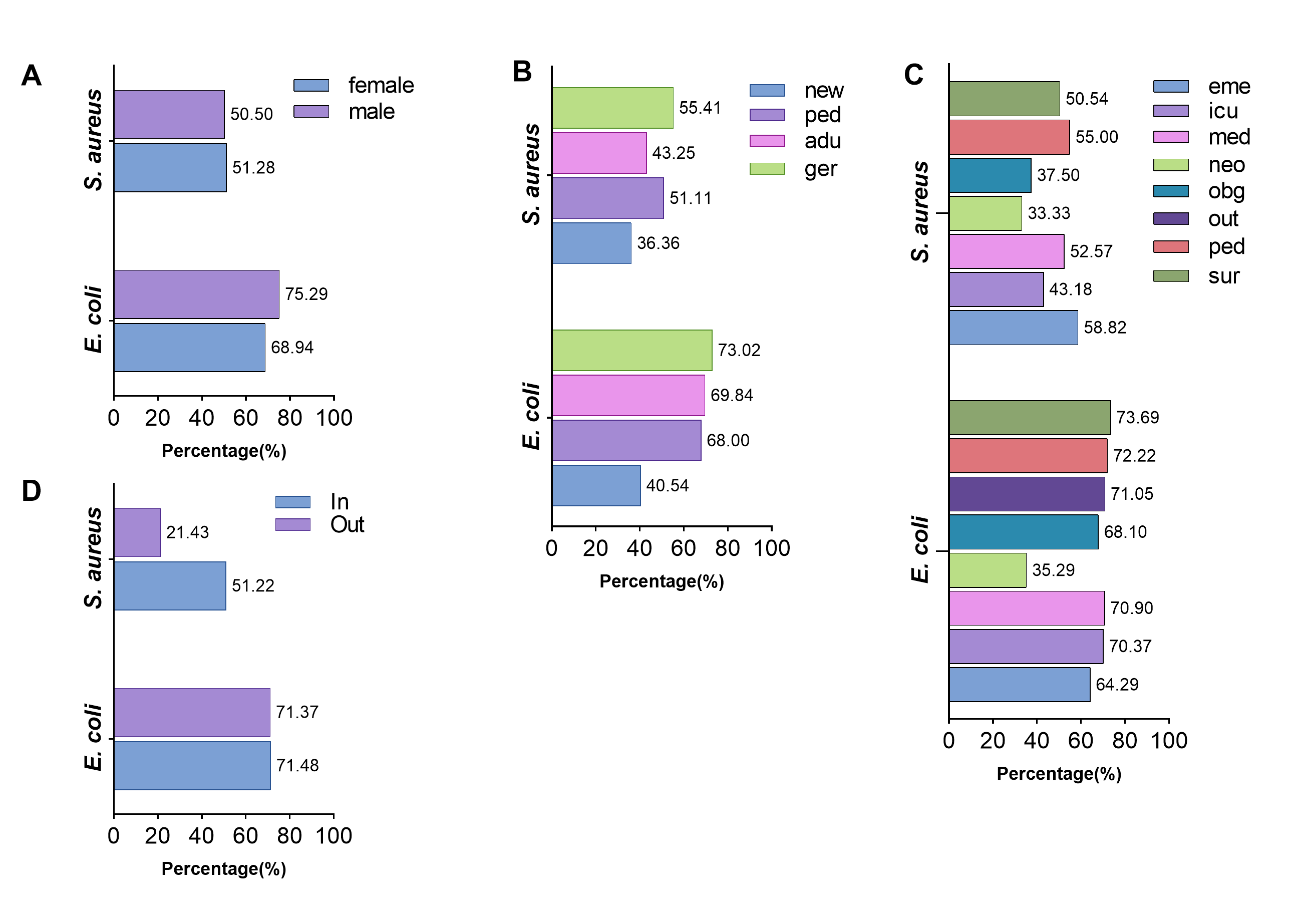
Subsequently, the AMR traits within the hospital were further elaborated. In short, the differences in resistance rates to most antimicrobial classes were not related to gender and inpatient or outpatient regardless of whether they were infected with E. coli or S. aureus (supplement Fig. 1 and 2, supplement Table 3 and 4). Furthermore, the isolates from neonate and neonatology demonstrated the lowest resistance rates to almost all antimicrobial classes (supplement Fig 3 and 4). Additionally, the resistance rates of isolates obtained from sputum, throat swabs, and urine are higher than those of other sampling positions (supplement Fig 5).
Trends in variation of resistance rates and prevalence
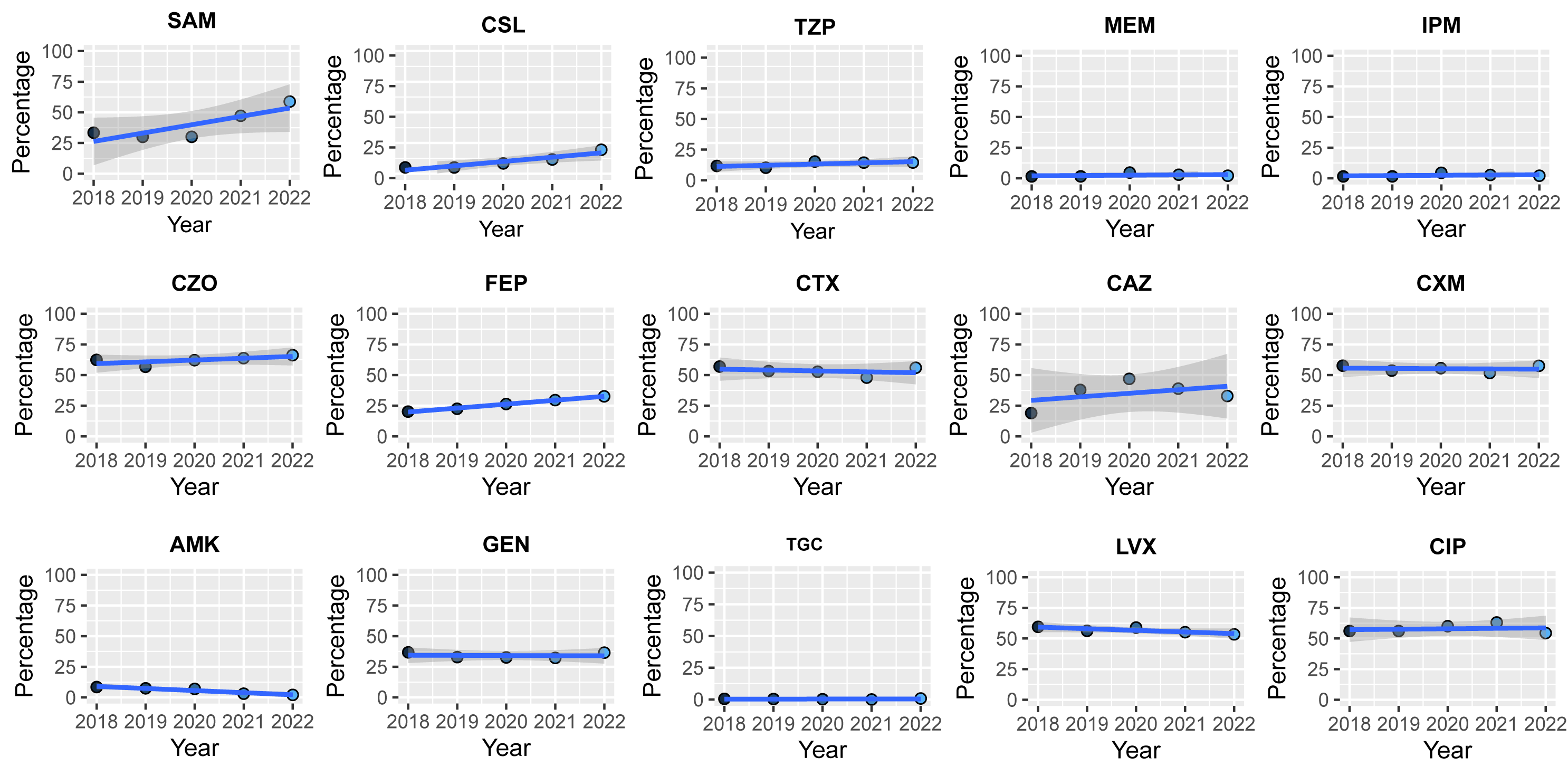
The clinical E. coli AMR dataset comprised 8 classes of continuous data spanning from 2018 to 2022. The linear trend test (2018-2022) revealed that resistance rates for six classes, namely β-Lactam combination agents, cephems, carbapenems, fluoroquinolones, folate pathway antagonists, and fosfomycin, exhibited linear trends. Of these, five classes showed an upward trend (R>0, P<0.05), while the class of folate pathway antagonist exhibited a downward trend (R=-0.072, P< 0.001, Fig. 2A). Notably, the change in resistance rate was relatively minor in magnitude (R<0.2). Furthermore, no discernible patterns in resistance prevalence were observed towards the 15 agents with continuous data (supplement Fig. 6).
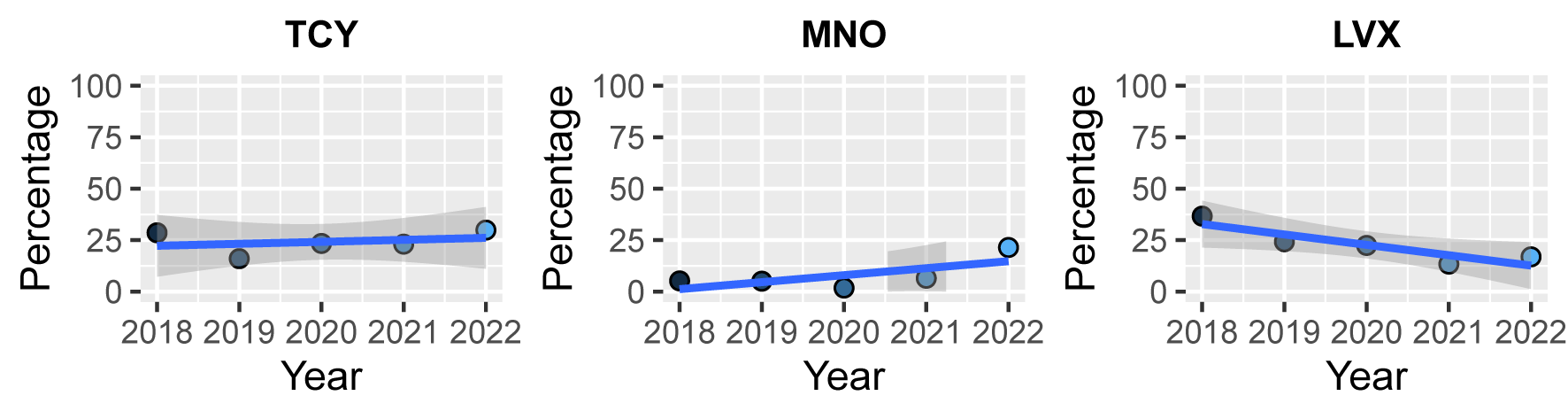
Linear trend tests of resistance rate to 11 classes with 5 years of continuous data, were conducted in clinical S. aureus isolates. There was a slight five-year downward trend observed in resistance rates to penicillinase-labile penicillins, penicillinase-stable penicillins, fluoroquinolones, lincosamides, and aminoglycosides, with the most significant decrease observed in resistance rates to fluoroquinolones (R=-0.177, P<0.001, Fig. 2B). In contrast, the resistance rates of S. aureus to folate pathway antagonists and tetracyclines showed a slight upward trend (R=0.215 and 0.281, P<0.001). No isolate resistant to glycopeptides and oxazolidinones was detected over the five years, and resistance rates to macrolides and ansamycins had no linear change trend (P=0.213 and 0.606). All classes of tetracyclines and fluoroquinolones included three agents in the clinical S. aureus AST schedule, and three agents of tetracycline, minocycline, and levofloxacin has continuous data frame, also showed no significant change (supplement Fig. 7).
The prevalence of extended-spectrum beta-lactamases (ESBL) positive E. coli and methicillin-resistant S. aureus (MRSA)
The ESBL-positive isolates account for 53.26% (1398/2526) of all tested clinical E. coli isolates (Fig. 3A). Meanwhile, the linear trend test showed no significant change in prevalence over the five years (P=0.136, Fig. 3B). The prevalence of clinical MRSA was 43.81% (435/993, Fig. 3C), and the prevalence showed a slight downward trend based on the linear trend test (R=-0.106, P< 0.001, Fig. 3D).
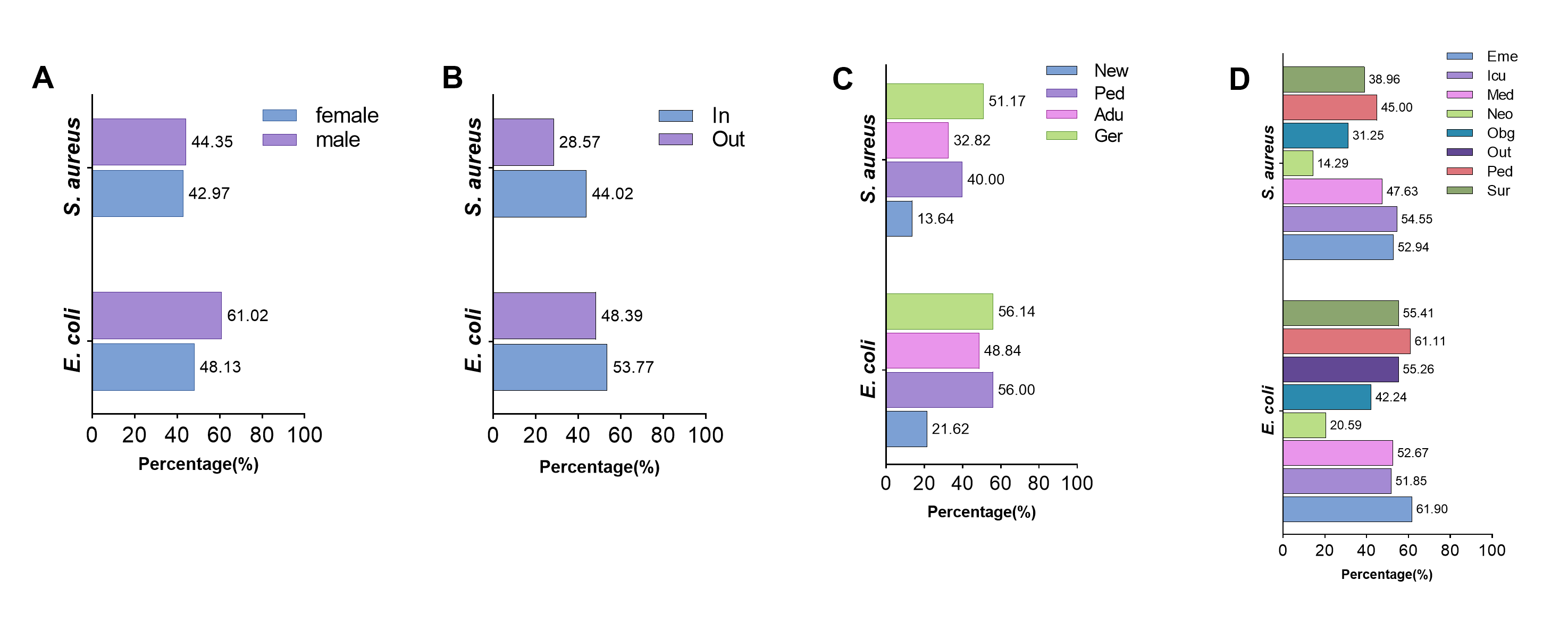
The characteristics of ESBL-positive E. coli and MRSA were further analyzed regarding gender, age, hospitalization status, and visiting department of hospital patients. There was a significant difference in the prevalence of ESBL-positive E. coli between male and female patients (61.02%/48.13%, χ²=41.91, P< 0.001, supplement Fig. 8), while there was no difference in that of MRSA (44.35%/42.97%, χ²=0.185, P=0.667). The prevalence of ESBL-positive E. coli (53.77%/48.39%, χ²=2.609, P=0.106) and MRSA (44.02%/28.57%, χ²=1.339, P=0.247, supplement Fig. 8) in inpatient was higher compared to outpatients, but the difference was not statistically significant. Out of the four age groups, the geriatric had the highest prevalence of ESBL-positive E. coli (56.14%) and MRSA (51.17%), whereas the neonate group had the lowest prevalence (21.62%/13.64%, supplement Fig. 8). In addition, the Department of Neonatology had the lowest prevalence of ESBL-positive E. coli (20.59%) and MRSA (14.29%).
Status and dynamics of MDR in clinical E. coli and S. aureus isolates
Out of the 2625 clinical E. coli isolates tested, 1877 (71.50%) were MDR isolates, which were resistant to three or more antimicrobial classes (Fig. 4A). None of the E. coli isolates were resistant to all 13 antimicrobial classes, and only one isolate was resistant to 12 classes, with no colistin resistance. There was an upward trend in the proportion of MDR isolates from 2018 to 2022, mainly those that are resistant to 9 to 12 classes (Fig. 4B and 4C). Notably, a considerable number of isolates have been detected that are resistant to 8 and 9 classes since 2021, and the emergence of isolates resistant to 10 and 11 classes started in the same year. Furthermore, the isolates resistant to 12 classes were detected in 2022.
The proportion of MDR S. aureus isolates was 50.80% (505/994), with none of the isolates exhibiting resistance to all 12 classes (Fig. 4D). Despite the lack of a discernible trend in the proportion of MDR S. aureus over the past five years (P=0.885), the highest proportion of MDR isolates was observed in 2022 (61.69%, Fig. 4E and 4F). Moreover, the isolates resistant to 8 and 9 antimicrobial classes were all identified in 2021 and 2022. The more details of MDR proportion of S. aureus in gender, age, hospitalization status, and visiting department of hospital patients was shown in supplement Fig. 9A and 9D.
The Poisson GLMMs were used to investigate the variation of AMR carriage (the total number of antimicrobial classes to which an isolate was phenotypically resistant) over the five years. The AMR carriages of E. coli in 2020, 2021, and 2022 were found to be higher compared to 2018 (odds ratio [OR]= 1.09, 1.31, and 1.59, P< 0.05), suggesting a deteriorating trend of E. coli MDR (Table 2). In contrast to E. coli, the AMR carriages of S. aureus in 2019, 2020, and 2021 were lower than that of 2018 (OR=0.83/0.91/0.78, P=0.003/0.147/0.001, Table 2), suggesting a decline in the MDR situation of S. aureus. Nevertheless, the slightly elevated AMR carriage of 2022 (OR=1.06, P=0.354) suggested that the issue of MDR should not be disregarded.
The AMR association between clinical and food animal isolates based on sole phenotype data
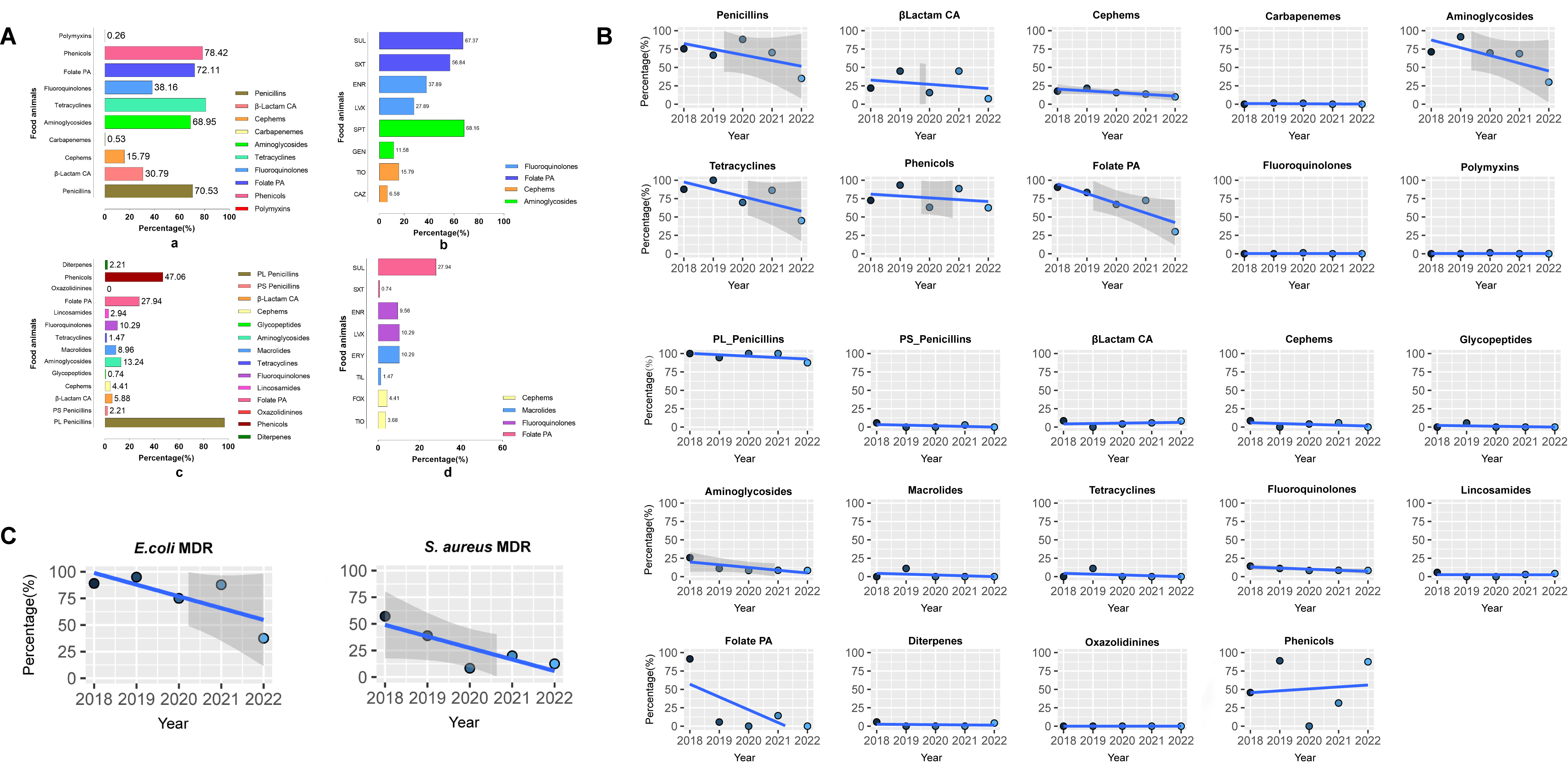
The AMR characteristics of E. coli and S. aureus isolates from food animals (or raw milk) are depicted in supplement Fig. 10A-C. The correlation coefficient of resistance rates was calculated to assess the AMR association between clinical and food-animal interfaces. The correlation coefficient heatmap for E. coli showed a strong negative correlation in the resistance rate of cephems between the clinical and food animal isolates (r=-1.000, P<0.001). No one-to-one correlation has been established between other phenotypic resistance rates, including the MDR rate. The analysis of the phenotype correlation between clinical resistance rates of S. aureus and isolates from food animals has also revealed the same phenomenon (Fig. 5). These results suggest that establishing connections at the human-animal interface through a single AMR phenotype is exceedingly challenging.
The shared AMR profiles between clinical and food animal interfaces
323 unique AMR profiles were identified in the clinical and food animal E. coli isolates including 250 profiles in clinical E. coli isolates, 29 profiles in food animal isolates and 44 shared by both interfaces (Fig. 6A).
Out of the 294 profiles in clinical E. coli isolates, the profiles contained six antimicrobial classes (antibiogram lengths 6) had the highest number of unique profiles (51 kinds), followed by antibiogram lengths 5 (47 kinds) and antibiogram lengths 4 (43 kinds, supplement Table 5, Fig. 6C). Notably, there was a noticeable rise in the diversity of AMR profiles, and the profiles with antibiogram lengths ranging from 7 to 12 were almost observed in 2020, 2021, and 2022 (Fig. 6D). Interestingly, each antibiogram length had at least one dominant AMR profile (supplement Table 5), including Pen-Fpa (78/293) and Pen-Flu (63/293) with antibiogram length 2; Pen-Cep-Flu (70/361) with antibiogram length 3; Pen-βLac-Cep-Flu (81/446) and Pen-Cep-Flu-Fpa (75/446) with antibiogram length 4; Pen-Cep-Ami-Flu-Fpa (63/371) and Pen-βLac-Cep-Flu-Fpa (56/371) with antibiogram length 5; Pen-βLac-Cep-Ami-Flu-Fpa (105/333) with antibiogram length 6; Pen-βLac-Cep-Ami-Flu-Fpa-Mon (40/170) with antibiogram length 7; Pen-βLac-Cep-Ami-Tet-Flu-Fpa-Mon (29/103) and Pen-βLac-Cep-Tet-Flu-Fpa-Mon-Phe (21/103) with antibiogram length 8; Pen-βLac-Cep-Ami-Tet-Flu-Fpa-Mon-Phe (28/64) with antibiogram length 9; Pen-βLac-Cep-Ami-Tet-Flu-Fpa-Mon-Fos-Phe (10/20) with antibiogram length 10; Pen-βLac-Cep-Ami-Tet-Flu-Fpa-Mon-Fos-Phe-Nit with antibiogram length 11 (4/8).
Out of the shared AMR profiles in E. coli, there were 7 AMR profiles in clinical scenarios in 2018, 2019, and 2020, consisting of four profiles with antibiogram length 1, two with antibiogram length 2, and one with an antibiogram length 3. In 2021, 24 new AMR profiles emerged, and an additional 13 profiles were observed in 2022. (supplement Table 6). In contrast, the AMR profiles of E. coli isolates from food animals were frequently observed in the initial three years (30/44), with 17 profiles in 2018, 6 in 2019, and 7 in 2020 (supplement Table 6). The results suggest that specific AMR profiles, particularly those with antibiogram length of three or more (MDR isolates), may exist earlier in the E. coli from food animals.
The S. aureus isolates from both clinics and food animals exhibited 141 AMR profiles, including 107 in clinical isolates, 22 in food animal isolates, and 12 shared by both interfaces (Fig. 6B). Among the 119 profiles of clinical isolates, both antibiogram length 3 and 4 exhibited 22 different AMR profiles, followed by antibiogram length 6 (19 kinds) and antibiogram length 5 (18 kinds, Fig. 6C). The AMR profiles in 2022 (68) and 2021 (54) were significantly higher compared to those in 2018 (33), 2019 (27), and 2020 (29). Additionally, there was an increase in the proportion of antibiogram length exceeding 3 in 2021 (68.52%, 37/54) and 2022 (83.83%, 57/68, supplement Table 6). Similarly, each antibiogram length of clinical S. aureus had one dominant AMR profile, namely PLPen-PSPen with antibiogram length 2 (99/204); PLPen-Mac-Lin with antibiogram length 3 (72/129); PLPen-PSPen-Mac-Lin with antibiogram length 4 (87/146); PLPen-PSPen-Mac-Flu-Lin with antibiogram length 5 (14/58); PLPen-PSPen-Ami-Mac-Flu-Lin with antibiogram length 7 (62/99); PLPen-PSPen-Ami-Mac-Tet-Flu-Lin with antibiogram length 7 (51/69, supplement Table 7).
The shared AMR profiles in S. aureus consist of 3 profiles with antibiogram length 3 and 1 with antibiogram length 4 (supplement Table 8). Interestingly, the four profiles also emerged in clinical isolates in 2021 or 2022 and were observed during the initial three years in food-animal isolates. This observation aligned with that of E. coli, which could potentially strengthen the validity of our conclusion.
Co-occurrence analysis discovered two apparent clusters in the resistant phenotype dataset of E. coli. One cluster was penicillins-β-Lactam_combination_agents-cephems- monobactams (Pen-βLac-Cep-Mon) in clinical isolates, and the other was aminoglycosides-tetracyclines-folate_pathway_antagomists (Ami-Tet-Fpa) in food animal isolates (P< 0.01, correlation coefficient > 0.5, Fig. 7A and B) was found in food animal isolates. The cluster of Ami-Tet-Fpa was detected in 10 AMR profiles of food animal isolates, 11 profiles of clinical isolates, and four profiles shared by both interfaces. As the monobactams class was not adopted in food animal E. coli AST, an alternative cluster of Pen-βLac-Cep was scanned at the human-animal interface and was detected in 125 AMR profiles of clinical isolates, eight profiles of food animal isolates, and six profiles shared by both interfaces (Table 3). Furthermore, 13 pairs of phenotypes were found in clinical E. coli isolates. Six co-occurrence pairs (3 pairs were same with clinic isolates) in food animal isolates were identified. Most of them (except Cep-Fos and Fos-Phe) were detected at human and food animal interfaces (Table 3). The results provide evidence of the potential association between human and food animal interfaces based on MDR phenotypes.
Co-occurrence analysis in the clinical S. aureus phenotypic dataset identified three distinct clusters of Ami-Mac-Lin (correlation coefficient > 0.75), Ami-Mac-Flu-Lin, and PSPen-Ami-Flu-Lin (correlation coefficient > 0.5, Fig. 7C). The three clusters were all presented in clinical and food animal AMR profiles (24.37%/0.29%, 9.24%/0.29%, 5.04%/0.57%, Table 3). In addition, three pairs of co-occurrence AMR phenotypes were identified in clinical S. aureus isolates, with only PLPen-PSPen being found in both clinical (34.35%) and food animal (0.86%) interfaces (Table 3). The AST for S. aureus from food animals incorporated three more classes of β-Lactam combination agents, cephems, and diterpenes. Thus, although three clusters of PSPen-βLac-Cep, PSPen-βLac-Cep-Mac and PSPen-βLac-Cep-Flu-Lin-Dit were identified in phenotypic dataset of food animal S. aureus isolates, only PSPen-Mac (adjusted from PSPen-βLac-Cep-Mac, 31.09%/0.29%) and PSPen-Flu-Lin (adjusted from PSPen-βLac-Cep-Flu-Lin-Dit, 10.08%/0.57%) can be used to determine their presence or absence in both interfaces (Fig. 7D, Table 3). Furthermore, eight pairs of co-occurrence AMR phenotypes were identified in the food-animal phenotypic dataset, and 6 of these pairs were observed in both clinical and food-animal interfaces. The results obtained from S. aureus also provided insights into the potential use of MDR as a cross-interface association assessment tool for G+ bacteria.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}