3.1. Nanoemulsion stability analysis:
The stability of nanoemulsion was observed in terms of their appearance and droplet size [38]. The samples were stored undisturbed for 15 days at 25 ºC for observation. On visual inspection, there was no creaming and flocculation in the nanoemulsion observed during the storage period. Additionally, the stability of the nanoemulsions was evaluated by droplet size measurement. A rapid increase in droplet size corresponds to an unstable nanoemulsion. Fig. 2 displays the droplet size of 1% SE and 1% PE on the 0th day and 15th day, respectively. The average droplet size observed in 1% SE was in the range of ~ 244 nm to 285 nm and ~ 164 nm to 297 nm in 1% PE. As evident from Fig. 2, the size of the maximum droplets of nanoemulsions was stable over a period of 15 days. The result indicated that the larger droplet size, greater than a micron has been separated from the emulsion.
3.2 Analysis of dissolved free CO2 concentration in nanoemulsions:
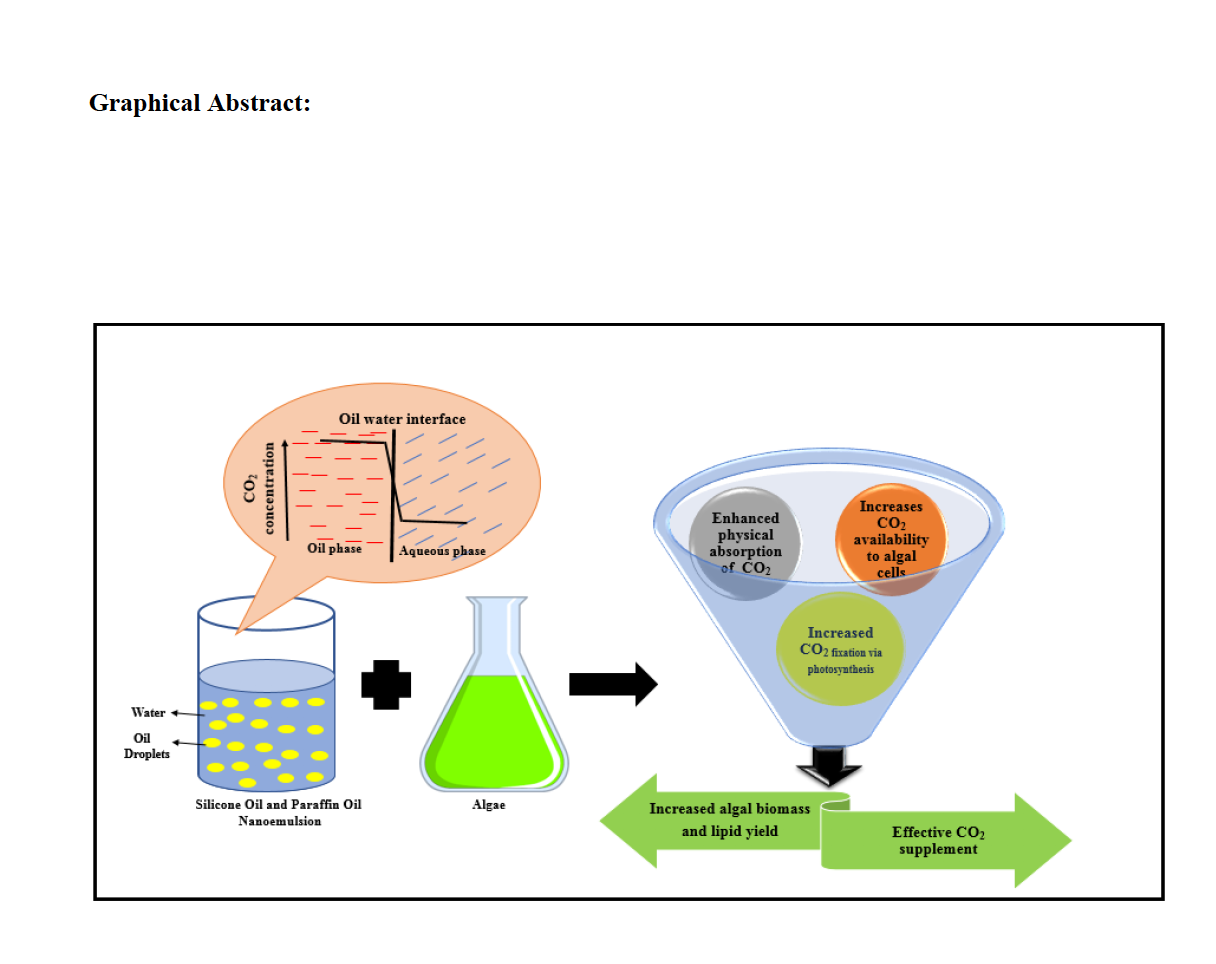
To determine the CO2 uptake efficiency in nanoemulsions, the CO2 absorption experiment was carried out using 1% SE, 1% PE and deionized water. In all cases, the DCC value increased with time and reached saturation within 1 hour. The results depicted that 1% SE retained the maximum amount of CO2 followed by 1% PE and DI water. The pH was dropped relatively fast in DI water than nanoemulsions with the addition of CO2 and the final DCC observed in DI water was 8.8 mg L-1, due to the absence of ions. On the other hand, in nanoemulsions pH dropped slowly and the final DCC observed was 14.1 mg L-1 in 1% SE and 13.2 mg L-1 in 1% PE which suggested that an additional amount of CO2 was retained in nanoemulsions compared with DI. The reason for increased CO2 concentration in 1% SE and 1% PE might be due to the presence of organic phase (i.e. oil) in the nanoemulsions which absorbed additional CO2 as the partial pressure of CO2 during sparging was higher. Based on the obtained result, it can be interpreted that 1% SE and 1% PE retained 60% and 50% higher free CO2 compared with DI water.
3.3. Growth of microalgae in nanoemulsions:
In the present study, 1% SE and 1% PE were prepared to test its potential in microalgal cultivation. Cell growth in formulated nanoemulsions was evaluated by the following parameters: optical density at 680 nm, pH, Chl-a, biomass yield, biomass productivity, and cell count.
Interestingly, for both the nanoemulsions, C. pyrenoidosa showed an increase in growth compared with conventional BG11 media (control). Results obtained from measuring OD 680, showed that~ 1.6 folds growth was observed in 1% SE supplemented growth medium compared with control on the 10th day. On the other hand, 1% PE growth medium gave an increment of ~ 1.8 fold in microalgal cell growth compared with control. In 1% SE and 1% PE supplemented microalgal cultivation systems, the accelerated growth phase was observed from day-1 compared with control. This prolonged growth phase lasted till the 10thday of the microalgal cultivation in nanoemulsions while in control lasted till the 8th day.
Gonçalveset al., [39] reported that increased CO2 concentration in the microalgal cultivation system extends the exponential phase. Araujo et al., [40] observed that the addition of CO2 during cultivating of diatom (Chaetoceros cf. wighamii) protracted the exponential phase and this phase of the life span of microalgae has the highest nutritional value when used for aquatic animals as feed. In the present study, the growth pattern including the duration of the prolonged growth phase was quite similar for both the nanoemulsions. The microalgal cell growth during the accelerated phase was much higher in nanoemulsion systems compared with control (Fig. 3).
The increase in microalgal growth in nanoemulsions was probably due to the increased CO2 captured by the nanoemulsions from the atmosphere compared with control as discussed in the section. 3.2. The higher CO2 content in nanoemulsions was supported by the pH measurement of nanoemulsions substituted microalgal cultures. The change in pH observed in control (~ 41%) was comparatively lower than the change in pH observed in 1% SE (~ 89%) and 1% PE (~ 90%). The increase in pH was observed in cultivation with 1% PE (~ 5.4 to 10.3) and 1% SE (~ 5.2 to 9.9) nanoemulsion, which was probably due to an increase in hydroxyl ion concentrations during uptake of bicarbonates and CO3-2. In the case of 1% PE substituted algal cultivation, the CO3-2 could be the dominant species which probably increased the pH value above 10. The higher pH change suggests that more microalgal cells were replicated in nanoemulsions substituted media compared with control as the rise in pH of cultivation media directly corresponds to the algal growth [41]. This study suggested that the 1% SE and 1% PE act as suitable alkalescent culture media which was appropriate for the growth of microalgae [42].
Furthermore, pigment synthesis in terms of Chl-a was investigated in C. pyrenoidosa cultivated in 1% SE, 1% PE and control as a function of culture time. It was found that 1% SE and 1% PE enhanced Chl-a pigment synthesis by 76% and 53% compared with control (Fig. 3). The increase in Chl-a concentrations in nanoemulsions cultivated biomass could be due to the sufficient availability of CO2 to the microalgal cells. Additionally, it might be possible that small oil droplets of nanoemulsion increase the residence time of light inside nanoemulsion leading to enhanced availability of light and enhanced pigment synthesis by microalgal cells (explained in 3.6). Besides, a slight decrease (~15%) in Chl-a content was observed in microalgal cells cultivated in 1% PE compared to 1% SE. This change is directly linked with the rise in pH during microalgal cultivation. Increased pH results in the volatilization of ammonia [43]. The volatilization of ammonia affects the nitrogen metabolism of algal cells and creates nitrogen limiting conditions, thereby impacting the chlorophyll content in microalgae [44]. Literature also suggests that the increase in CO2 concentration leads to acidification of the algal cultivation system. This results in the replacement of Mg+2 with H+ and the formation of pheophytin instead of chlorophyll [45]. Salehi et al.,[46] studied the effect of n-alkane on Chlorella vulgaris and reported that the existence of hydrocarbons in the medium increases the permeability of the cell wall, resulting in the accumulation of hydrocarbons inside the algal cell. This might be the reason for the inconsistency between chlorophyll concentration and cell concentration in the case of 1% PE because n-alkanes are the major portion of paraffin oil.
Hence, overall it can be summarized that 1% SE and 1% PE nanoemulsions act as active CO2 carriers which promote microalgal growth and Chl-a synthesis comparative with conventional growth media (BG11).
The biomass yield of C. pyrenoidosa obtained in both the nanoemulsions was higher compared with control (~ 1.03 g L-1) (Fig. 4). Comparing both the nanoemulsions, the biomass yield of C. pyrenoidosa was higher in 1% PE (~ 3.20 g L-1) than 1% SE (~ 2.75 g L-1). Table 1, highlights the biomass yield of Chlorella sp. obtained using different growth media by various researchers. Out of these, Chi et al. [47] obtained maximum biomass yield of Chlorella sp. was about 1.10 g L-1 in Bold Basal media. Chandra et al. [48] reported maximum biomass yield of Chlorella minutissima was 1.84 g L-1 while cultivating in a modified CHU-13 medium by varying different abiotic factors. Additionally, Choix et al. [49] studied biomass yield of Chlorella vulgaris U162, Chlorella sp., Scenedesmus obliquus U169, and Scenedesmus sp. by filtered Tequila vinasses and bio digested as cultivation media. The maximum biomass yield was recorded as 2.30 g L−1 obtained from Chlorella sp. cultured in Tequila vinasses. The biomass concentration of Chlorella sorokiniana was recorded 1.00 g L-1 while using CO2 from flue gas as a carbon source [50]. Wong, [51] used five different media to grow Chlorella sp. and got a maximum biomass concentration of 1.42 g L-1 using Bold Basal Media. In another such study, Prajapati et al. [35] found the biomass yield 0.98 g L-1using Tap media. To efficiently deliver CO2 to algal cells, Zheng et al. [52] used a membrane-based system and found an increase in biomass concentration and the maximum biomass yield achieved was 1.77 g L-1 with CO2 loading. In another study, Spirulina sp. LEB 18. was cultivated in the Zarrouk medium with a mixture of diethanolamine (1.64 mmol L−1) and potassium carbonate (0.41 mmol L−1). The combination of diethanolamine and potassium carbonate enhanced the dissolved inorganic carbon concentration in the cultivation medium resulting in an increased biomass concentration of 2.10 g L-1 [53]. Interestingly, the biomass yield in 1% SE and 1% PE was higher compared to all the reported studies so far, signifying the importance of this cultivation media.
Table 1: Comparison of biomass yield of Chlorella sp. in different cultivation media
|
Growth Medium
|
Biomass yield (g L-1)
|
References
|
|
Bold Basal Media
|
1.10
|
[47]
|
|
Modified CHU-13
|
1.84
|
[48]
|
|
Tequila vinasses
|
2.30
|
[49]
|
|
BG11+ CO2 from flue gas
|
1.00
|
[50]
|
|
Bold Basal, Modified BG-11
|
1.42 ± 0.01, 0.90±0.01
|
[51]
|
|
Carbonate Based Media at 0.5 and 0.7 loading concentration
|
1.63 ±0.10 and 1.77 ± 0.02
|
[11]
|
|
Zarrouk medium + mixture of diethanolamine and potassium carbonate
|
2.10
|
[53]
|
|
Urea +K2HPO4+ MgSO4·7H2O and Ammonium ferric citrate
|
1.37
|
[98]
|
|
Tap-water
|
0.98 ± 0.11
|
[35]
|
|
1% SE and 1% PE amended with BG-11
|
2.75 ± 0.07and 3.20 ± 0.07
|
Present study
|
Correspondingly, the biomass productivity was evaluated after the 12th day. The biomass productivity of C. pyrenoidosa in 1% PE and 1% SE was 0.26 and 0.22 g L-1d-1 respectively. The biomass productivity of C. pyrenoidosa was found 0.08 g L-1d-1 in control which was lower than 1% PE and 1% SE (Table 2). Yu et al. [54] obtained biomass productivity (0.24 g L-1d-1) of Chlorella vulgaris in a mixotrophic mode of cultivation. Chen et al. [55] reported 0.31 g L−1 d−1 biomass productivity of Chlorella sp. C2 in the presence of nutrient-rich ash and flue gas. Similarly, the periodic addition of monoethanolamine and CO2 during the cultivation of Chlorella fusca LEB 111 yield biomass productivity of 0.15 g L-1d-1 [13]. Chlorella sp.T12 was cultivated in four different cultivation media (NSIII, Chu No.10, BG11, and BBM) and obtained the highest biomass productivity (0.20 g L-1d-1 ) in BBM media [56]. Furthermore, the productivity of Chlorella fusca LEB 111 was reported 0.14 g L-1 d-1 cultivated in the presence of a magnetic field [57]. In another study, Chlorella vulgaris was cultured in seawater media and obtained 0.13 g L-1 d-1 biomass productivity in presence of the following: 2% CO2, light intensity 10000 Lux, 0.3% Walne nutrient concentration and, 7 days of cultivation period [58]. Similarly, Shabani et al. [59] cultivated Chlorella vulgaris in various levels of salinity and CO2 concentration and found biomass productivity of 0.09 g L-1 d-1 in natural water at 10% CO2 on the 8th day. The marine microalgal strain Nannochloropsis sp. was cultivated in presence of CO2 delivered through semipermeable membranes along with chemical solvent with biomass productivity of 0.10 g L-1 d-1 [60]. Biomass productivity of C. pyrenoidosa obtained in both the nanoemulsions in the present study was significantly higher than the reported results.
Table 2: Comparison of biomass productivities of Chlorella sp. in different cultivation media
|
Growth Medium
|
Biomass Productivity (g L-1d-1)
|
References
|
|
BG11 + Monoethanolamine+ CO2
|
0.15
|
[13]
|
|
BG11 + CO2 + Acetic Acid
|
0.24
|
[54]
|
|
BPPA2
|
0.31
|
[55]
|
|
Bold’s Basal Media
|
0.20
|
[56]
|
|
BG11
|
0.14
|
[57]
|
|
Seawater media
|
0.13
|
[58]
|
|
Bold’s Basal Media + Natural water + CO2
|
0.09
|
[59]
|
|
1% SE and 1% PE amended with BG-11
|
0.22 ± 0.07 and 0.26 ± 0.07
|
Present study
|
The better CO2 absorption in the organic phase of nanoemulsions could have resulted in high microalgal biomass productivity. A similar study was performed by Sawdon and Peng, [61] using perfluorocarbon emulsion for the cultivation of C. vulgaris in tubular photobioreactor. They observed an increase in microalgal cell concentration 4-folds, because of efficient CO2 delivery to microalgal cells by perfluorocarbon emulsion. Apart from this, silicone oil had also been studied in the delivery of respiratory gases (CO2 and O2) in microbial cultivation [62], mammalian cell culturing [63], and in human retinal treatment [64].
To further cross confirm the above results, direct cell counting was performed for observing the microalgal cell population in 1% SE, 1% PE, and control on the 12th day of the experiment using a cell counter. Result showed that 1% PE had the maximum cell number (3.0 ± 0.21 x 106 cells mL-1), followed by 1% SE (2.4 ± 0.30 x 106 cells mL-1) and control (1.34 ± 0.09 x 106 cells mL-1). In a similar such study where Chlorella vulgaris was cultivated with 8% CO2 concentration, a cell count of 1.3 × 107 cells mL-1 on 10th days [65]. In another study during the cultivation time of 20 days, Chlorella sorokiniana gave cell density of 5.82 ± 0.03 x 106 cells mL-1 [66]. However, the exact comparison cannot be made as the strain type, cultivation media, and growth conditions vary in the reported studies. In the present study, the obtained cell numbers were in agreement with the cell growth and biomass yield obtained from 1% PE and 1% SE nanoemulsions. Hence, from the present study, it was concluded that the 1% SE and 1% PE have successfully enhanced the potential of BG11 media by acting as an efficient growth supplement for microalgal cells. However, as mentioned in section 3.2, the DCC of 1% SE was higher than the 1% PE but the microalgal growth, cell count and biomass yield obtained was higher in 1% PE supplemented microalgal cultivation. This could be due to the chemical composition of paraffin oil and the most probable explanations were highlighted in section 3.6.
3.4. MTT, SYTOX® Green Staining:
The viability of cell indicates the reproductive capacity of cells and also represent the physiological state which involves the production of enzymes like oxidoreductase, etc. [67]. In the case of microalgal and cyanobacterial cultures, tetrazolium compounds have been used for viability assessment [68]. Hence in this study, the viability testing of microalgal cells grown in 1% SE and 1% PE emulsion has been conducted on the 8th day of cultivation using tetrazolium dye i.e. MTT via MTT assay. The sensitivity of the MTT assay is based on the ability of succinate dehydrogenase, microalgal mitochondrial enzyme, to convert MTT to a water-insoluble formazan dye in viable cells. The viability of cells is based on the intensity of purple color, which shows that darker the color, the more the viability in cells. In present study, the OD570 measured for microalgal cells cultivated in 1% PE was higher than 1% SE and control (Fig. 5a). This represents that microalgal cells were more viable in both the nanoemulsions compared with control. Additionally, the obtained results conclude that 1% PE and 1% SE nanoemulsions were non-toxic to the microalgal cells. Further, using SYTOX® Green and fluorescence microscopy, a clear distinction between live and dead cells was observed. SYTOX® Green stain is impermeable to the cell wall but can stain the nucleic acid of cells when the cell wall has been damaged. The damaged cell wall allows the diffusion of SYTOX® Green stain inside the cells with the compromised cell membrane and binds with the nucleic acid of the cells to generate green fluorescence. The observations confirmed that most of the microalgal cells cultivated in 1% SE and 1% PE had intact chloroplasts and membranes (Fig.5b) as evident from emitted red fluorescence due to autofluorescence of inherent chlorophyll found in microalgal cells [69]. Only a small fraction of cells were stained by the green fluorescent dye SYTOX® Green. On the other hand, the control culture showed relatively large patches of green emissions under a fluorescence microscope. Hence, from the results obtained from both MTT and SYTOX® Green staining, it can be concluded that the maximum number of microalgal cells cultivated in nanoemulsions were healthy and viable on the 8th day of cultivation leading to higher cell density and biomass yield.
3.5. Characterization
3.5.1. Bio-compositional analysis
The bio-compositional analysis was performed to evaluate the quantitative changes in macromolecules (protein, lipids, and carbohydrates) of harvested microalgal biomass cultivated in 1% SE and 1% PE compared to control (Table 3). It has been reported that during the stationary stage of cell growth comes, nutrient depletion occurs which results in a shift of carbon flow to storage compounds like carbohydrates and lipids [70]. An elevated CO2 concentration in microalgal cells could trigger the Calvin cycle, resulting in the conversion of available CO2 into biomolecule syntheses like carbohydrates and lipids. The biosynthesis pathway of lipids and carbohydrates compete for the same carbon source, but carbohydrate synthesis requires less energy than lipids [70]. The carbohydrate content of microalgal biomass recovered from 1% SE and 1% PE was ~18.9% and 17.2% respectively, which was higher than control. Rosa et al. [71] reported that the delivery of CO2 through monoethanolamine enhances the biomass production and carbohydrate content (28.2% w/w) of Spirulina sp. LEB 18. Ji et al. [72] observed an increase in carbohydrate contents of Scenedesmus obliquus were 22.3%, 23.9%, and 23.6% at 5%, 10%, and 14.1% CO2, respectively. The finding suggests that an increase in the availability of CO2 to microalgal cells in cultivation media leads to enhanced carbohydrate synthesis inside microalgal cells [72]. However, the composition and metabolism of carbohydrates vary from species to species. Therefore, to select microalgal species with efficient carbohydrate productivity is highly recommended for biofuel production [73].
Table 3: Biomass (biochemical) composition of C. pyrenoidosain terms of lipid, carbohydrate, and protein content. The values were stated as the mean ± SD.
|
Biochemical composition
|
Control
|
1% SE
|
1% PE
|
|
Carbohydrates (%)
|
13.6 ± 0.56
|
18.9±0.28
|
17.2 ±0.56
|
|
Proteins (%)
|
51.75±0.77
|
53.75 ± 0.07
|
47.1±0.14
|
|
Lipids (%)
|
18.05 ± 0.35
|
23.6±0.84
|
26.8±0.84
|
Additionally, an increase in lipid content in algal biomass obtained from 1% SE (23.60 % ± 0.84) and 1% PE (26.80 % ± 0.84) with respect to control (18.05% ± 0.35). Interestingly, in both the nanoemulsions, the highest lipid content was observed in microalgal biomass harvested from 1% PE. Ji et al. [72] reported that lipid contents of Scenedesmus obliquus were increased with an increase in CO2 concentration and were evaluated as 19.80%, 21.50%, and 22.80% at 5%, 10%, and 14.1% CO2 respectively. Wang et al. [74] also described that Chaetoceros muelleri showed increased lipid accumulation and biomass production on exposure to a 10% CO2 concentration. Similarly, Tang et al. [75] reported an increase in lipid content and microalgal growth in presence of 10% CO2 in Scenedesmus obliquus and Chlorella pyrenoidosa. The results obtained were in agreement with the reported studies.
In order to investigate the relationship between biomass productivity and lipid content, lipid productivity was calculated. The biomass of Chlorella pyrenoidosa obtained from 1% PE had the highest lipid productivity (69.6 mg L-1 d-1) followed by 1% SE harvested biomass (51.9 mg L-1 d-1) compared with BG11 grown biomass (14.4 mg L-1 d-1). The lipid productivity of C. pyrenoidosa was enhanced by 4.8 and 3.6 folds in 1% PE and 1% SE compared with BG11. The obtained productivity was higher than 49.1 mg L-1 d-1 lipid productivity of Chlorella minutissima attained by Arora et al. [76] by optimizing nitrogen and phosphorous concentration. Ordog et al. [77] observed 60 mg L-1 d-1 lipid productivity at 30 °C and 2% nitrogen in Chlorella sp. Jiang et al., [78] cultivated Chlorella sorokiniana in anaerobically digested effluent from kitchen waste with seawater and obtained lipid productivity of 19.00 mg L-1 d-1. However, lipid productivity is the strain-specific response that also depends on cultivation conditions and biomass productivity. Consequently, the variation in lipid productivity of Chlorella sp. might be observed in the literature. Hence from the present study, it can be concluded that nanoemulsions positivly affected the lipid productivity of Chlorella pyrenoidosa. The obtained results showed a positive relationship between biomass productivity and lipid content.
The protein content was slightly enhanced in 1% SE cultivated microalgal cells (53.75% ± 0.07) compared to control, indicating they can be used as aquaculture feedstock, which desires protein content in the range of 35 to 60 wt% [79]. The protein content of microalgal biomass harvested from 1% PE (~ 47%) decreased compared with microalgal biomass of control (~ 51%) and 1% SE (~ 53%). The high concentration of CO2 affects the N metabolism of algal cells indirectly. As an outcome, the assimilation of nitrogen inside algal cells increases which create nitrogen limiting condition [80]. Therefore, the concentration of protein, as well as chlorophyll decreases. This claim supports our experimental results obtained by 1% PE supplemented microalgal cultivation. Enhanced lipid and carbohydrate content obtained from microalgal biomass recovered from nanoemulsions suggests better prospects in various applications.
3.5.2. FTIR Analysis
The microalgal samples were observed using Fourier Transform Infrared (FTIR) technique (Table 4). The identification of peak is based on a comparison of the bands from recorded FTIR spectra of microalgae with reference literature [81]. The FTIR transmittance of the C. pyrenoidosa showed the presence of Si-O, P=O, C-OH, -CO, -COOH, functional groups (Fig. 6). IR spectra of C. pyrenoidosa showed higher intensity of lipid and carbohydrates functional groups from algal biomass of 1% SE and 1% PE. The strong peak at 1652-1653 cm-1, 1266 cm-1, 1089 cm-1 representing amide I, protein, and nucleic acid proteins were observed in microalgal biomass recovered from 1% SE. The bands at 1200–950 cm-1 have shown absorption strength in biomass of both i.e., 1% SE and 1% PE which directs the presence of carbohydrate content compared to control. Similarly, Ansari et al. [82] reported the presence of a strong peak in the range of 1000-1200 cm-1 in Chlorella sp. which depicts carbohydrate potential. Meng and Kassim [83] reported similar findings of an increase in band intensity in the region of carbohydrate in presence of elevated CO2 concentration in microalgae. The intense peak was found in the region 1000-1150 and 2200-3000 cm-1, which is less evident in control cells. This represents the accumulation of carbohydrates and lipids of microalgal cells harvested from nanoemulsions. Two regions are commonly used for the assessment of lipid content. One is at 1740 cm-1, conveying stretching of the ester bond, and other between 2800-3000 cm-1 having a methyl and methylene group [84]. The strong absorption bands observed at 1456, 1744, 2852, 2922, 2924 cm-1 clearly shows the presence of membrane lipids and lipids representing CH2, CH3, C=O groups in biomass obtained from1% SE and 1% PE compared with control. Correspondingly, Zawar et al. [85] described an absorption peak at 2926 cm-1 in Chlorella sorokiniana which signifies CH2 stretching of lipids. While, peaks at ~1266 cm-1 represents carbohydrates, protein, DNA, and RNA respectively [86]. It was clearly visible from the FTIR spectra of nanoemulsions recovered biomass of C. pyrenoidosa possess fingerprints of lipids, carbohydrates, and proteins. FTIR spectroscopy appears as a viable analytical tool for revalidating the analytical data on microalgal biomass composition. The present findings are in agreement with the earlier studies for numerous microalgal species including C. vulgaris, C. reinhardtii, etc [85,86].
Table 4: Monitoring macromolecular changes in C. pyrenoidosa by FTIR analysis
|
Wavenumber range (cm-1)
|
Assignments
|
Functional groups
|
Peak (cm-1)
|
|
1064-880
|
-------
|
Carbohydrate
|
965
|
|
1090-1030
|
P=O or
Si-O
|
Nucleic acids
|
1089
|
|
1150-1000
|
C-O/v Si-O
|
Polysaccharides/Siloxane (Carbohydrate peak/siloxane shoulder at 1200 cm-1)
|
1072
|
|
1263
|
C-O
|
Ester
|
1260
|
|
1275
|
C-O-H
|
Carbohydrates, proteins, DNA, and RNA
|
1266
|
|
1398-1370
|
CH3, CH2, C-O
|
Proteins, Carboxylic Groups
|
1394
|
|
1456-1450
|
CH2, CH3
|
Lipid, Protein
|
1456
|
|
1550-1640
|
N-H bending
|
Amide
|
1558
|
|
1655-1638
|
C=O
|
Protein (Amide I)
|
1652-1653
|
|
1745-1734
|
C=O of esters
|
Membrane Lipids, Fatty acids
|
1744
|
|
2875-2850
|
CH2, CH3
|
Lipids
|
2852
|
|
2930-2920
|
CH2
|
Lipids
|
2922,2924
|
3.5.3 Microscopic Analysis:
The biomass of C. pyrenoidosa obtained on a day 12 was subjected to FESEM analysis to investigate the morphological variation in the cell structure. Fig. 7 shows the FESEM images of C. pyrenoidosa biomass harvested from BG11, 1% SE, and 1% PE. From these images, it is evident that the cells were spherical and the size of the microalgal cells in control was 6 ± 2 µm which was quite similar to the cell size in 1% SE and 1% PE. In the case of nanoemulsions (Fig. 7 b and 7c), the cells retained their basic shape, but the edges of the cell wall were not similar to cells of control. Besides cells grown in nanoemulsions appeared morphologically intact with no cell damage. However, the microalgal cells grown in nanoemulsions were found in aggregation and displayed presence of a surface coating. This can be attributed to the release of an extracellular polymeric substance (EPS) when cells are grown in nanoemulsion. The EPS is a sticky substance released by algal cells and is composed of polysaccharides and proteins [87]. The presence of enhanced CO2 in 1% SE and 1% PE nanoemulsions might have triggered the microalgal cells to release this substance. It has been reported that the elevated CO2 concentration stimulates the metabolic carbon flux of algal cells and in order to maintain carbon balance, algae excrete organics like EPS [88]. The compositional analysis of obtained microalgal biomass also showed that carbohydrate content was elevated in nanoemulsions substituted cultivation which can be correlated with EPS secretions. A study suggested that secretion of EPS is known to be associated with carbohydrate content [89]. Though the present study did not quantify EPS, enhanced carbohydrate contents and aggregation could be attributed to this. As observed from the FESEM images, the intracellular adhesion behavior of microalgal cells during nanoemulsion based cultivation opens the possibility to explore biofilm-based systems. The intracellular adhesion and cellular colonization are the most influential parameter for biofilm formation [90]. The cellular colonization restricts temporary immobilization of cells and ensures cellular bridging which increases the cell density and promotes thickening of biofilm which might help in gaining biomass yield and value-added products [91].
3.6. Other possible mechanisms for improved algal growth in nanoemulsions:
It should be noted that the nanoemulsions used in the present study possess higher solubility of CO2 compared with conventional growth media. However, it was not obvious that the higher solubility of CO2 was the only reason for increased microalgal growth in nanoemulsions substituted algal growth media. Based on the literature several other factors could have resulted in greater microalgal biomass productivity in conjugation with nanoemulsions. Some of the possible mechanisms are summarized as follows:
- In our preliminary investigations, a higher concentration of oil (1%, 2%, 3%, 4%, and 5%) was used to observe the growth of pyrenoidosa. The high concentration of oil was supposed to enhance dissolved CO2 concentration in nanoemulsion, but microalgal cell concentration declined at a higher concentration of oil like 3%, 4%, 5% [22]. The results suggested that an increase in silicone oil concentration leads to a decrease in microalgal growth. Hence, the inference was that diminished microalgal cell growth was probably due to the low penetration of light in the media at a higher concentration of oil in the microalgal culture. Therefore, a lower concentration was used for the cultivation of microalgae in the present study.
- Sawdon and Peng, [61] have reported the use of perfluorocarbon (PFC) to improve algal photosynthesis by decreasing dissolved oxygen in a tubular photobioreactor. The mechanism proposed by the authors was the absorption of oxygen by the PFC suspended in the culture which was produced by the algal cells during photosynthesis. In the proposed study also, the organic compounds possess higher solubility of oxygen [92] compared to water or the conventional growth media. Therefore, these organic solvents (silicone and paraffin oil) might help to decrease oxygen content in the continuous phase of nanoemulsions which indirectly promotes photosynthesis. However, more controlled experiments need to be carried out to observe the impact of reduced oxygen in the algal cultivation systems.
- Earlier studies have been reported that nanosuspension can also enhance the mass transfer of solutes [93]. In a quiescent fluid, diffusion is the main mechanism for mass transfer. For the case of nanosuspensions, in addition to diffusion, due to the Brownian motion of the suspended particles/droplets local disturbance velocity fields are also generated [94]. The resultant microscale convection of the fluid can enhance the mass transfer rates of solutes in a suspension. In the present study also, the droplets of oil could be contributing to improved mass transfer of nutrients, dissolved gases, etc. from the bulk growth media to the algal cells.
- Scattering and absorption of light by the suspended particles in a suspension reduces the availability of light in a media and can lower the rate of photochemical reactions [95]. However, the nature of suspended particles is transparent and used in a small volume, we observed that the residence time of light inside the suspension can increase (manuscript under preparation). Both the oils used in the present work were used in a small volume and transparent, which suggests that there is an increase in the residence time of light inside nanoemulsions. This might improve the light availability in the nanoemulsions supplemented media and the microalgal cells can efficiently utilize light to carry out photosynthesis.
- Paraffin components were found as a stimulant for algal growth at lower concentrations [97]. In general, the lower carbon content oil phase can work as a source of carbon to the algal cells. This could be the possible reason for the phenomenal increase in microalgal growth in 1% PE along with enhanced CO2 availability for microalgal growth.





{kind=link}