To our best knowledge, this study represents the inaugural application of MR analysis to explore the relationship between lipids, lipid-modifying pharmacological interventions, and IPF. Our results offer genetic corroboration for the proposition that PCSK9 inhibitors may elevate the risk of IPF. Notably, no evidence was found to suggest that lipid traits have a causal association with IPF risk, which implies that the mechanism by which PCSK9 inhibitors influence IPF risk may be distinct from its effects on lipid metabolism. Additionally, the study uncovered preliminary evidence hinting at a potential positive association between NPC1L1 inhibition and a higher IPF risk. The discovery of these results illuminates the possible adverse effects linked to the use of lipid-modifying drugs and offers an understanding of potential risk factors that may be investigated for IPF management.
Although the current evidence did not support serum lipid traits as causal risk factors for IPF, the result should be interpreted with caution. Firstly, lipids are categorized into four primary groups, including glycerides, fatty acids, non-glycerides, and lipoproteins. The plasma lipid traits predominantly featured in this MR study pertain to the lipoprotein class. However, extant literature suggests that dysregulation of phospholipid and sphingolipid metabolism is a more substantial contributor to IPF pathophysiology[9, 43, 44]. A lipidomics analysis has revealed that alterations in the plasma lipid profile of IPF patients are predominantly within the glycerophospholipid class. Among the 159 glycerolipids examined, 30 exhibited significant disparities between the IPF group and controls[44]. Moreover, the lung, being a lipid-rich organ, engages in intricate lipid metabolic processes. Compared with blood lipids, alveolar lipid levels may provide a more precise reflection of metabolic disturbances. Despite a paucity of research directly comparing lipidomics profiles between bronchoalveolar lavage fluid and blood samples from IPF individuals, the work of Marissa O 'Callaghan et al. offers some valuable insights. This study observed a marked increase in total lipid content within the lung tissue of IPF individuals relative to controls[45]. In addition, they also assessed pulmonary fat attenuation volume through chest CT images (CTPFAV). The median CTPFAV in IPF was greater compared to controls, however, there was no association observed with serum lipids and body mass index[45]. These findings imply that extracellular lipids within the lung may have a closer relationship with IPF than blood lipid traits. Future MR studies could explore the bidirectional causality between intrapulmonary lipid traits and IPF.
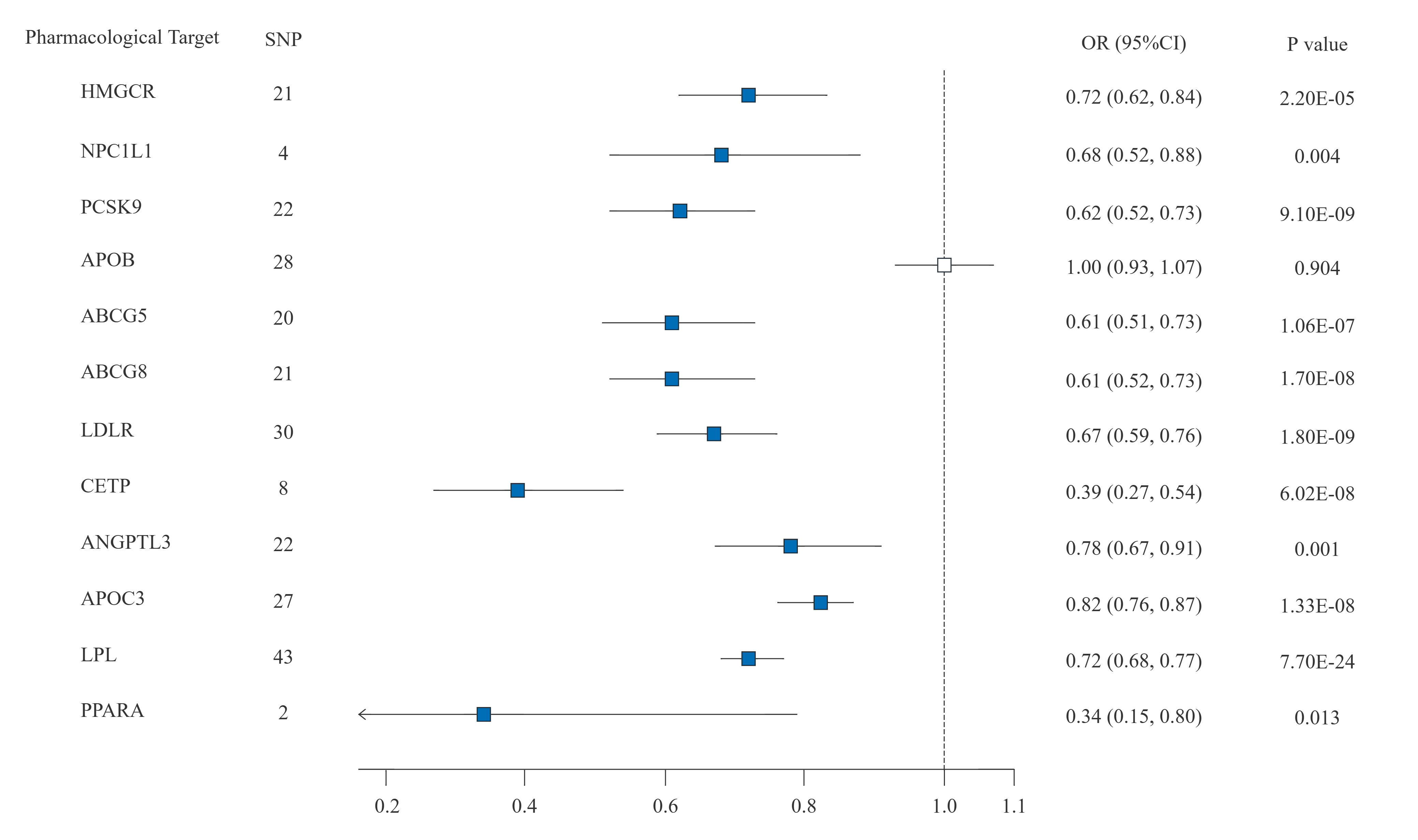
The MR study we conducted revealed a notable association between PCSK9 inhibitors and IPF. Contrary to manifesting as a protective effect, this association was characterized as a risk factor, which contradicts findings from previous studies. PCSK9, an enzyme, is essential for regulating cholesterol metabolism and maintaining cardiovascular health. Recent studies have shown that the PCSK9 gene may be implicated in the fibrotic processes of the liver, heart, kidney, and other organs. A study by Stefania Grimaudo revealed that increasing PCSK9 expression in male mice led to faster progression of liver fibrosis[46]. This research also described a protective role for the PCSK9 loss-of-function mutation against the progression of liver fibrosis. Subsequent research has indicated that anti-PCSK9 treatment may hold the potential to mitigate liver fibrosis by modulating the AMPK/mTOR/ULK1 signaling pathway, thus reducing hypoxia-induced autophagy in hepatocytes[47]. Another in vitro experiment showed that increasing PCSK9 levels could enhance the transition of cardiac fibroblasts into myofibroblasts, impacting fibrosis post-myocardial infarction[48]. Similar protective effects were also observed in renal fibrosis. Danyu Wu et al. developed a therapeutic vaccine targeting PCSK9. The results indicated that this vaccine could ameliorate kidney fibrosis by controlling fatty acid β-oxidation[49].
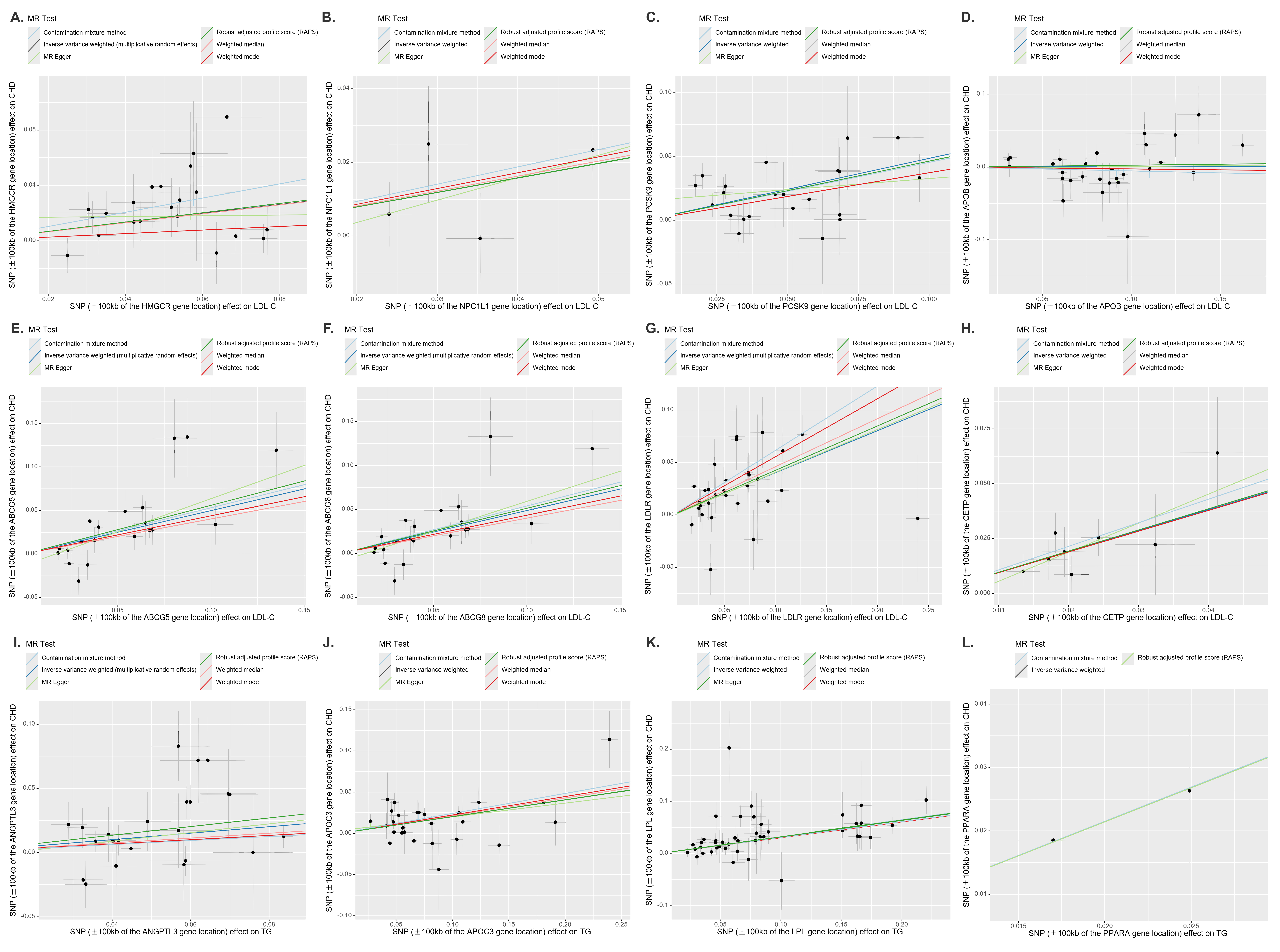
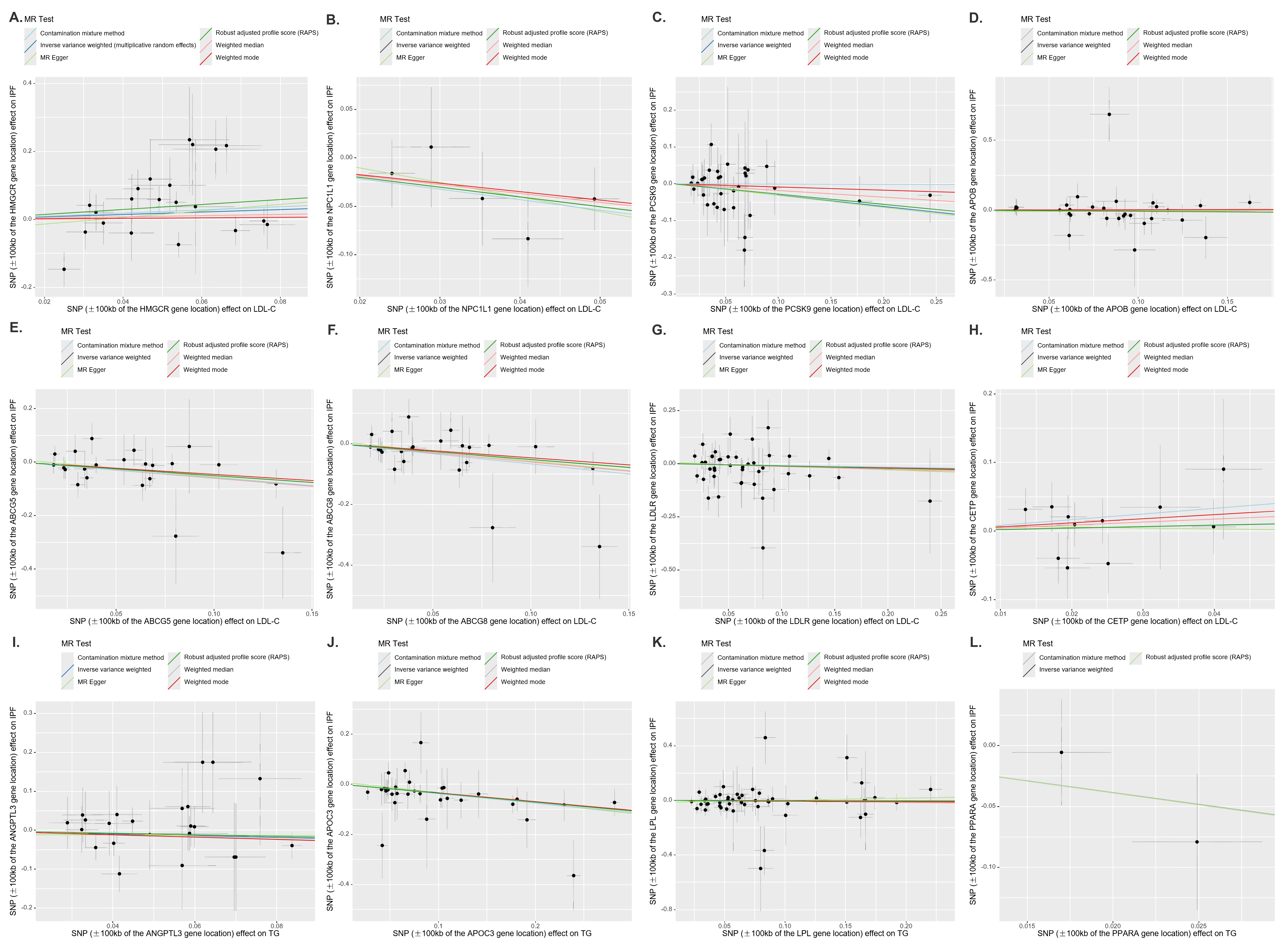
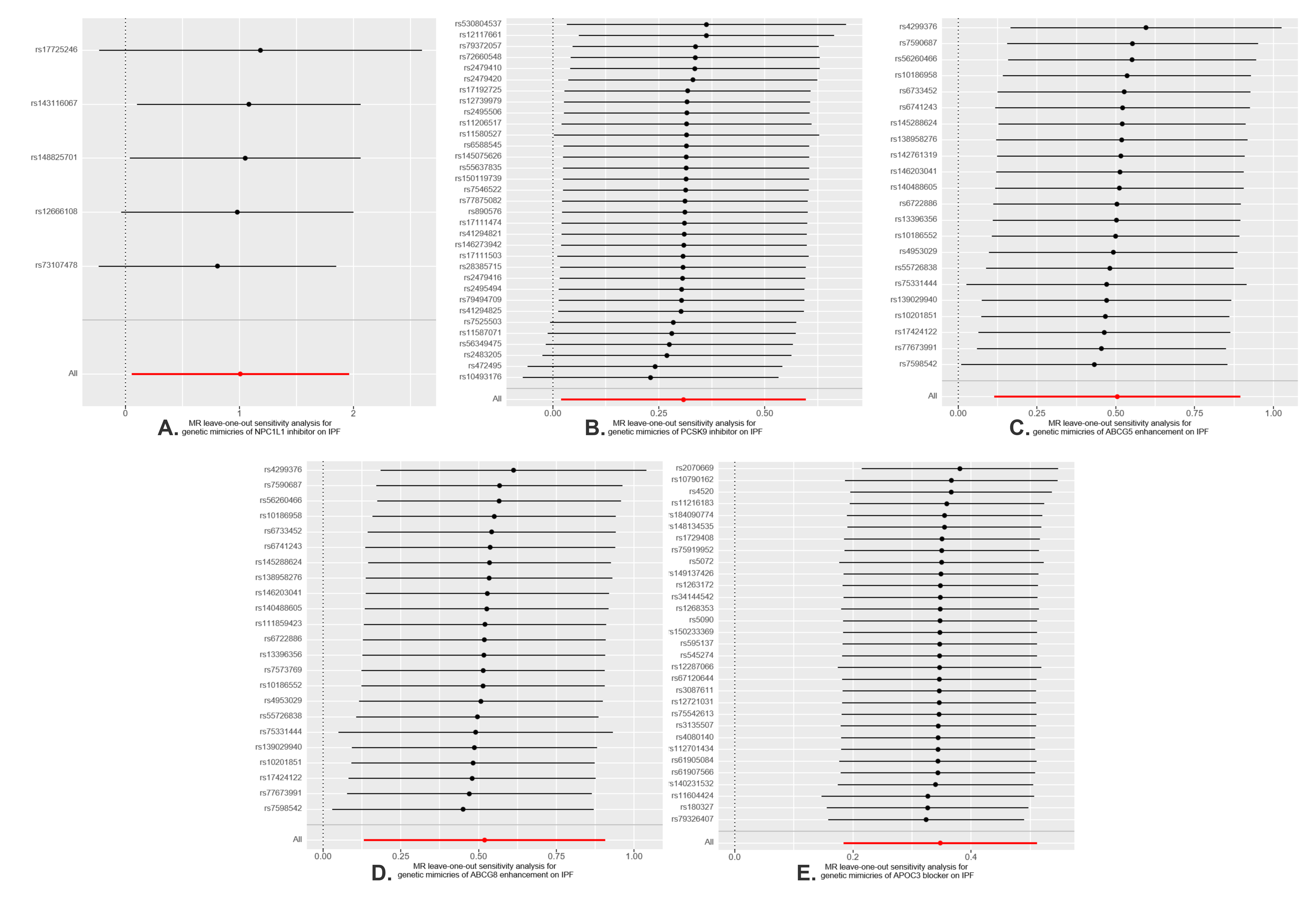
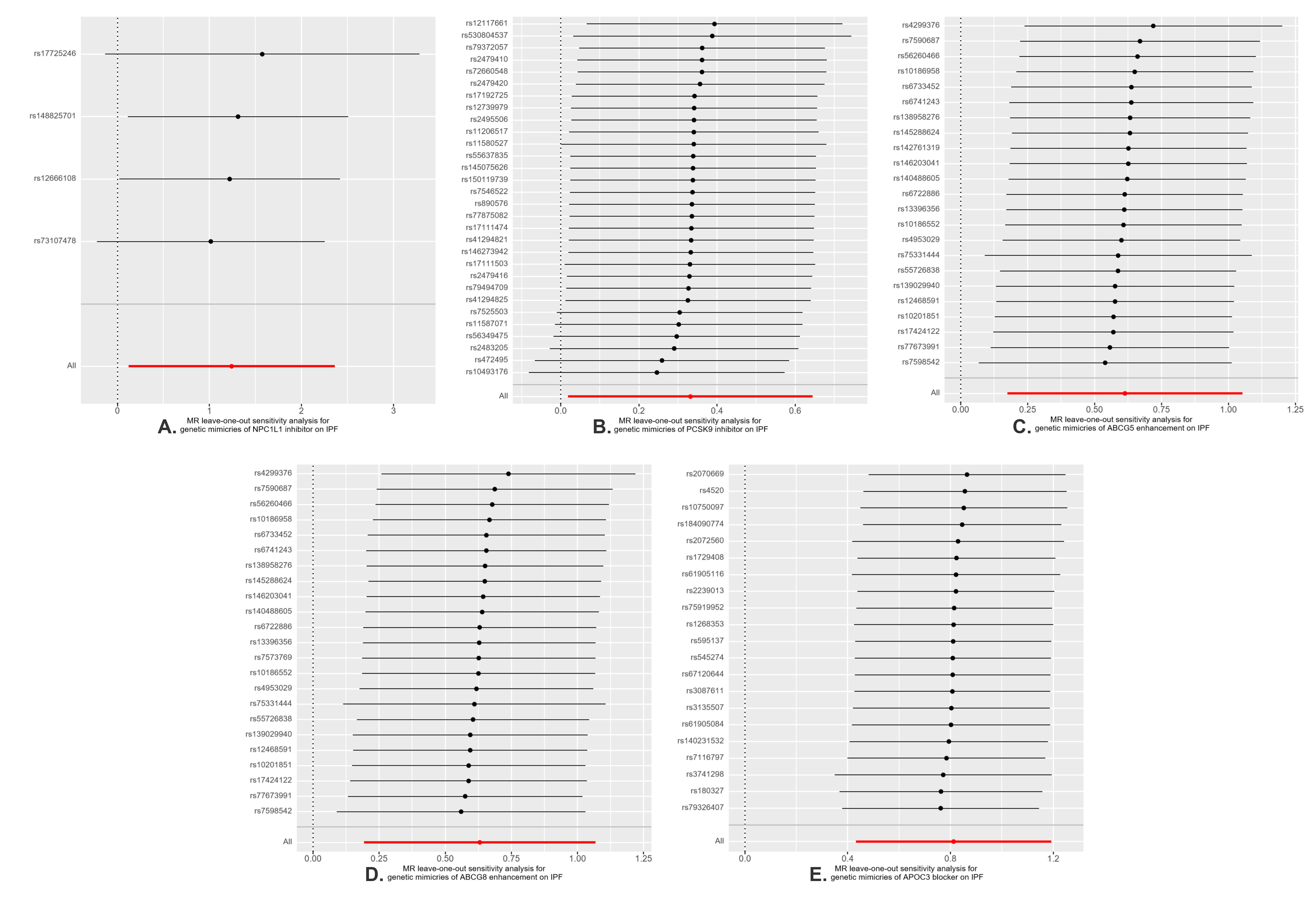
We postulate that several reasons may account for the inconsistent results observed. Firstly, the pathogenesis of IPF is highly complex, and the role of PCSK9 is also not singular. Previous research has concentrated on the anti-fibrotic effect of PCSK9 inhibitors in different organs through autophagy and oxidative regulation, rather than its impact on lipid levels. The balance between the anti-fibrotic effects of PCSK9 inhibitors through similar pathways or the pro-fibrotic effect through other pathways in lung tissue requires further investigation. Secondly, the expression levels of target genes can vary in different tissues, as evidenced by our SMR analysis of PCSK9 gene expression across different tissues. This variability can influence the therapeutic efficacy of drugs in a tissue-specific manner. Thirdly, drug-target Mendelian randomization analysis primarily models impact of gene inhibitors or blockers on outcomes using SNPs within the gene-specific action range (± 100kb). It provides information on the trends of connections rather than the tangible therapeutic advantages of drugs in practical situations. Numerous factors, such as drug dosage, when the drug is given, how inter-individuals metabolize drugs, and how well drugs attach to their intended targets, need to be further considered. Consequently, further RCTs are essential to confirm these observational findings.
Our findings also imply that there is a potential for increased IPF risk with exposure to NPC1L1 inhibitors, although the SMR analysis only suggested a near-positive result. Ezetimibe is known as the primary drug for NPC1L1 inhibitors, which are responsible for facilitating the absorption of dietary cholesterol by NPC1L1 protein [27]. To date, limited research has examined the influence of ezetimibe on the onset of IPF. Chanho Lee et al. conducted a retrospective study of the medical records across three different hospitals and discovered that individuals with IPF who consistently used ezetimibe had lower all-cause mortality and lung function decline rates[50]. They also found that ezetimibe could prevent mice from developing bleomycin-induced pulmonary fibrosis by suppressing mTORC1 activity in vitro study. The enhancement of autophagy in mouse lung fibroblasts mediates the anti-fibrotic effect of ezetimibe, rather than through lipid-lowering properties[50]. There may exist differences in the pathogenesis between drug-induced pulmonary fibrosis and IPF, which could account for the discrepancies between our MR findings and the in vitro results.
This MR research also has some limitations. Firstly, despite the numerous sensitivity analyses conducted that have reinforced the reliability of our results, the possibility of horizontal pleiotropy cannot be entirely dismissed. Secondly, the sample size of eQTL data is limited, and there are no eligible eQTLs for NPC1L1, ABCG5, ABCG8, and APOC3 in hepatic and pulmonary tissues, which are the primary organs involved in lipid metabolism. This limitation may result in an underestimation of the role these genes play in the pathogenesis of IPF. Thirdly, the initial GWAS data did not categorize by specific subtypes (such as the extent of LDL-C elevation). Consequently, this study was constrained from performing a stratified analysis, an approach that should be contemplated with the availability of more specific datasets in the future. Lastly, we lack a validation cohort because other GWAS data for IPF may have sample overlap with lipid exposure. It also must be acknowledged that the data presented in our study pertains to individuals of European descent. When extrapolating these results to different ethnic groups, caution should be exercised. Additional research encompassing a diverse array of populations is necessary.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}