Chronic activation of SST neurons of the prelimbic cortex has anxiolytic- and antidepressant-like effects on motivated behavior in male but not female mice.
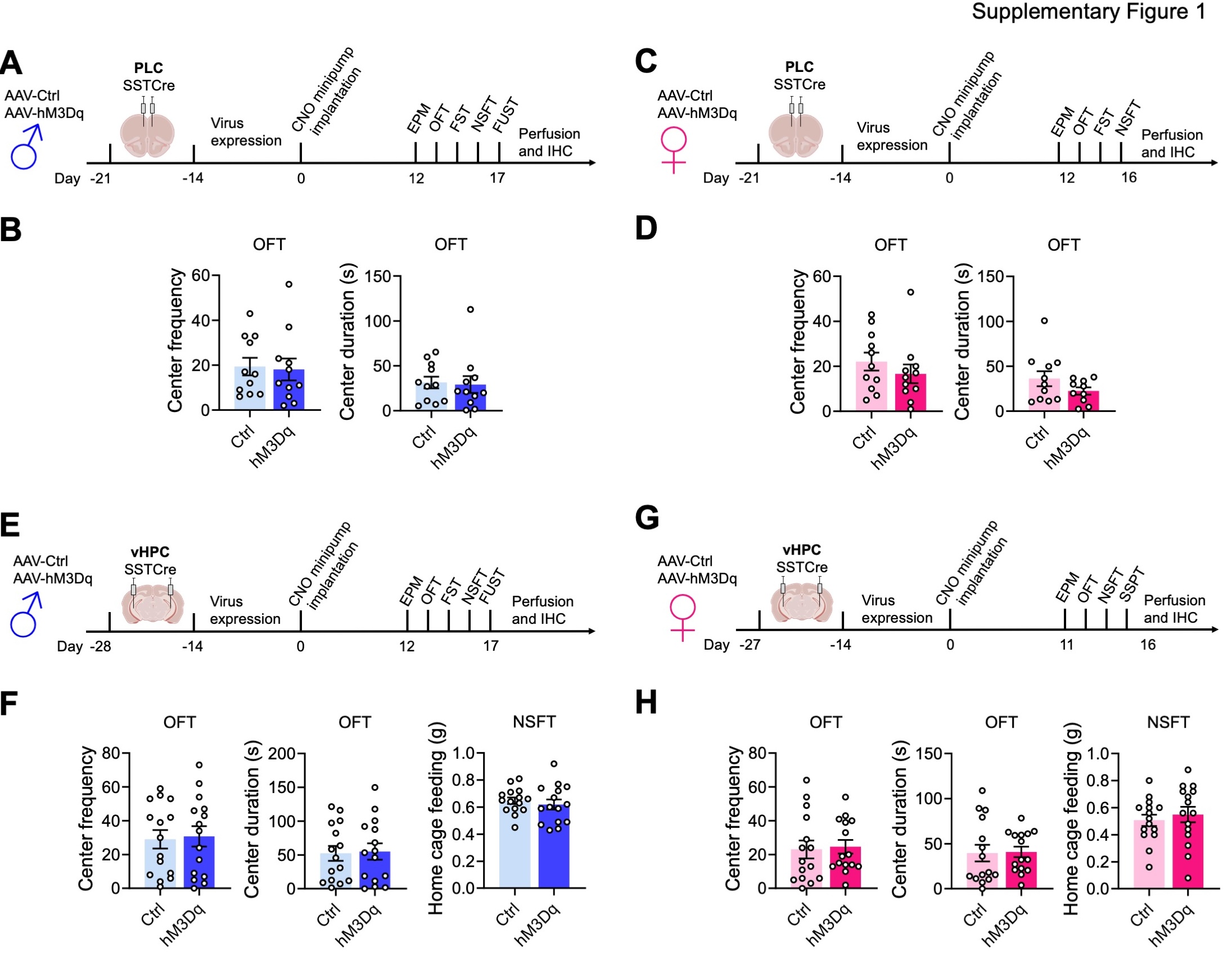
We previously showed that global disinhibition of SST neurons in SSTCre:γ2f/f mice results in behavior that mimics the effects of antidepressant drug treatment in both male and female mice. 24. SST neurons of SSTCre:γ2f/f mice are hyperexcitable due to loss of GABAergic inhibitory synaptic input and a significant increase in input resistance 24. Moreover, SST neurons are present throughout the brain as well as in diverse peripheral tissues, and the mutation is induced during embryonic development. This raised the question of whether the behavioral phenotype of SSTCre:γ2f/f mice can be reproduced by increasing the excitability of SST neurons in specific brain regions in adulthood. To address this question, we injected the prelimbic cortex (PLC) of male SSTCre mice (8–10 weeks of age) with a Cre-dependent AAV-hM3Dq-mCherry vector (hM3Dq) or a Cre-dependent AAV-mCherry control vector (Ctrl) and, after allowing three weeks for virus expression, implanted all animals with clozapine-N-oxide (CNO)-releasing osmotic minipumps, regardless of virus injected. We aimed to mimic the hyperexcitability of SST neurons observed in SSTCre:γ2f/f mice in AAV-hM3Dq-manipulated SSTCre mice with a low but constant dose of CNO (~ 0.22 mg/kg/h). Behavioral testing of male mice was initiated 12 days after minipump insertion and continuous CNO exposure using the test sequence depicted in Fig. 1A and sFigure 1A. We used the EPM, OFT, and NSFT to assess changes in locomotion and negatively regulated, anxiety-related behavior under low and comparatively high acute stress conditions, respectively. In addition, we used the FUST to assess changes in positively regulated motivated behavior and hedonic drive in male mice. The sniffing of urine in this test is associated with dopamine release in the nucleus accumbens, indicating it is rewarding 35. Lastly, these initial experiments included the FST as a test traditionally used to assess antidepressant drug activity in rodents. The behavior of PLC-injected male mice in the EPM (Fig. 1B) and OFT (sFigure. 1B) was unaffected by hM3Dq/CNO-mediated chronic activation of SST cells, indicating unaltered locomotion and anxiety-related behavior under low acute stress conditions. However, in the FST and NSFT, hM3Dq injected mice showed a reduced immobility score and a reduced latency to feed while in the FUST, they showed an increased time sniffing urine compared to empty vector controls, Thus, chronic activation of SST neurons in the PLC of male mice resulted in antidepressant drug-like changes in both positively and negatively regulated forms of motivated behavior.
Next, we analogously tested the effects of PLC SST neuron activation in female mice (Fig. 1C, D, sFigure 1C, D). The behavior in the EPM and OFT was unaffected as in males (Fig. 1D, sFigure 1D). However, female mice differed from males in that activation of SST cells in the PLC had no effects in the NSFT and FST. This finding was unexpected because SSTCre:γ2f/f mice with disinhibited SST neurons exhibit anxiolytic and antidepressant-like changes in behavior independent of sex 36. The data suggested that GABAergic control of motivated behavior in the PLC is male-specific and predicted that GABAergic control of motivated behavior in female mice is controlled by a distinct brain substrate.
Chronic activation of SST neurons in the ventral hippocampus has anxiolytic and antidepressant-like effects on motivated behavior in female but not male mice.
In search for a substrate for SST neuron-mediated changes in motivated behavior of female mice, we next focused on the ventral hippocampus (vHPC) as a second major brain region implicated in the top-down control of motivated behavior (Fig. 1E–H, sFigure 1E–H). Interestingly, we found that the sex-specific effects of hM3Dq/CNO-mediated activation of SST neurons in the vHPC were opposite to those in the PLC, as indicated by the lack of an effect of SST neuron activation in the EPM, FST, NSFT, and FUST of male mice (Fig. 1E, F), and robust anxiolytic-like changes in the EPM and NSFT of female mice (Fig. 1G, H). As a substitute for the FUST and FST, we subjected the vHPC-manipulated female mice to a Sucrose Splash Test. Grooming behavior in this test is accompanied by dopamine release in the nucleus accumbens 34, indicating that the behavior is rewarding similar to sniffing female urine for male mice. However, the behavior of hM3Dq/CNO manipulated female mice in the SSPT was unaffected in this experiment (sFigure 1H). In retrospect, this is consistent with recent evidence that dopamine release and grooming behavior in this test is selectively stimulated after chronically stressful experiences 34, as we confirmed further below. Notably, the anxiolytic-like effect of SST neuron activation in the vHPC of females was seen both under low-stress (EPM) and comparatively high-stress conditions (NSFT), whereas in PLC-manipulated male mice it was only seen under high-stress conditions (NSFT). Behavior in the OFT was unaffected, independent of sex and brain region manipulated (sFigure 1). Moreover, home cage feeding of food-deprived vHPC-manipulated animals was unaffected indicating that the behavioral changes in the NSFT were not due to an altered feeding drive. Collectively, these experiments indicated a striking sex-specific dissociation in the top-down control of antidepressant- and anxiolytic-like motivated behavior by prelimbic and hippocampal GABAergic microcircuits.
SST neurons of the PLC and vHPC mediate resilience to chronic stress-induced defects in motivated behavior in male and female mice, respectively.
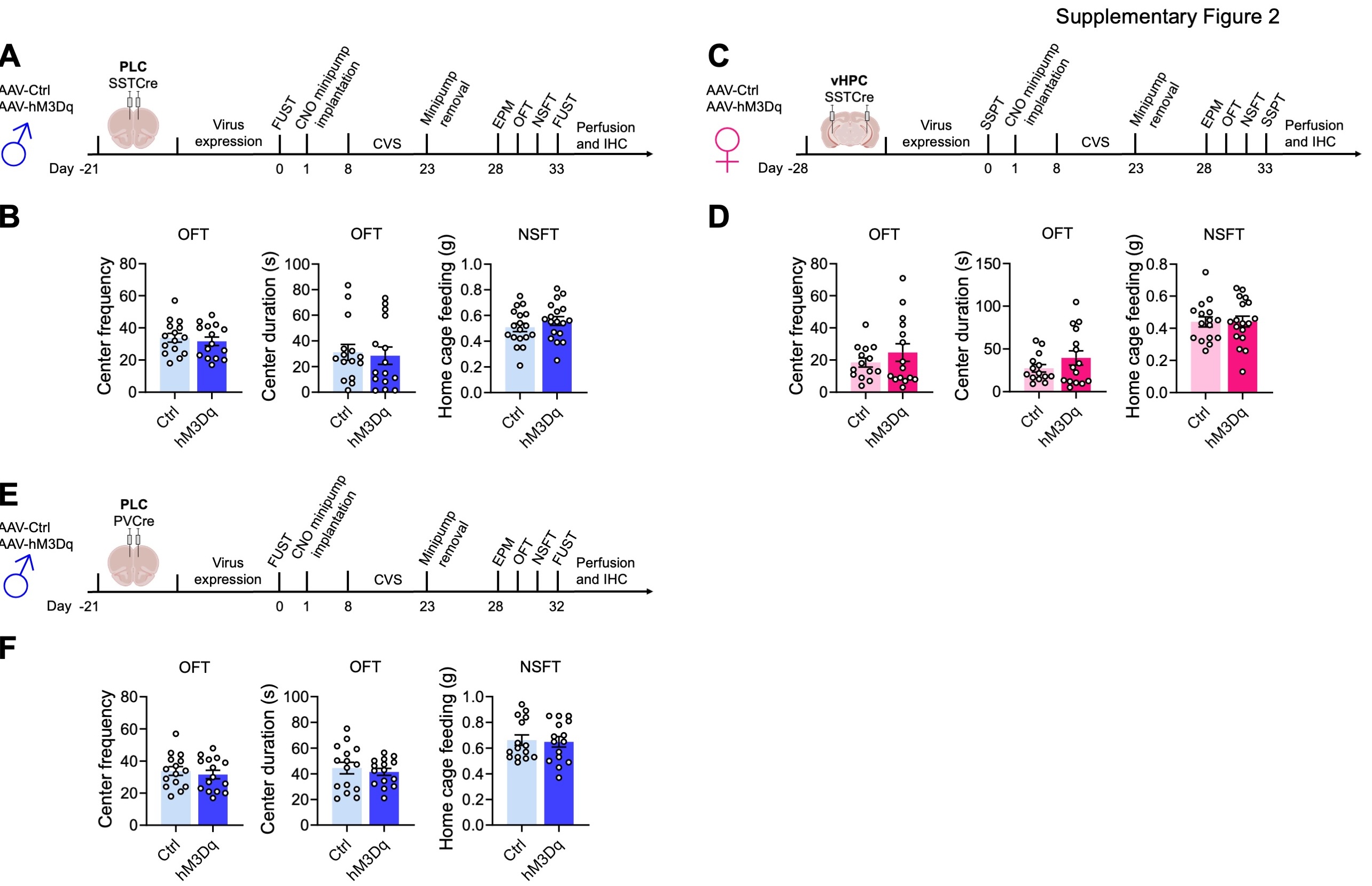
We next asked whether activation of SST neurons in the PLC of male mice is sufficient to confer resilience to chronic stress-induced defects in motivated behavior. Virus-transduced mice were allowed to recover and subjected to an initial FUST to assess baseline behavior, followed by CNO minipump implantation, 15 days of CVS, minipump removal, and a battery of tests assessing negatively and positively motivated behavior including a second FUST (Fig. 2A). The weight of mice during the three weeks of CVS was measured to assess chronic stress effects on whole-body physiology (Fig. 2B). Interestingly, CVS resulted in a time-dependent body weight reduction in control but not hMD3q-injected mice, suggesting whole-body physiological stress protection. Repeated testing in the FUST before and after CVS revealed an overall stress effect independent of the vector injected, as well as an interaction between CVS and the type of vector injected. CNO reversed the CVS effect in hM3Dq- but not control vector-manipulated mice (Fig. 2C). Analyses after CVS exposure further showed that hM3Dq-mediated activation of SST neurons had anxiolytic-like effects with regard to open arm entries and open arm duration in the EPM, and latency to feed in the NSFT. Collectively, these data indicated that SST neurons of the PLC were sufficient to mediate resilience to CVS-induced defects in both positively and negatively regulated behavior of male mice.
Next, we similarly tested whether SST neurons of the vHPC confer stress resilience to female mice (Fig. 2D). In contrast, to males, CVS failed to affect the weight of female mice independent of DREADD manipulation (Fig. 2E). Despite this qualitative sex difference in the stress effect, comparing the behavior in the SSPT before and after CVS revealed a significant CVS effect independent of virus (Fig. 2F, first panel). Post hoc analyses showed that this effect was significant in control vector- but not hM3Dq vector-injected mice. As in PLC-manipulated males, hM3Dq-injected female mice showed an anxiolytic effect in the NSFT (Fig. 2F, fourth panel). Therefore, SST neurons of the vHPC mediate resilience to CVS-induced defects in positively and negatively regulated behavior in female mice. Notably, SST neuron activation in the vHPC of females had no effect on % open arm times and open arm entries in the EPM (Fig. 2F, second and third panel), in contrast to SST neuron activation in the PLC of males (Fig. 2C), and the behavior in the OFT and home cage feeding of food-deprived mice was unaffected, independent of sex, brain region and interneuron type (sFigure 2A–F). These experiments suggest that the brain regions that mediate sex-specific antidepressant-like behavioral effects in the absence of stress also confer resilience to the detrimental behavioral consequences of chronic stress exposure. In male mice, GABAergic PLC-mediated stress resilience extends to stress-induced weight loss, whereas in female mice this parameter could not be addressed due to the absence of a significant stress effect.
Chronic activation of parvalbumin-positive interneurons in the PLC has detrimental effects in male mice.
Unlike the pro-resilience function of SST neurons in the PLC of male mice, increased activity of parvalbumin (PV)-positive interneurons may have detrimental effects, as shown by others for the mPFC of female and hippocampus of male mice 37, 38. To assess the function of PV neurons in the PLC of male mice, we here assessed the behavioral consequences of chemogenetic PV neuron activation in CVS-exposed mice. Comparing the female urine sniffing times before and after CVS revealed a significant stress effect independent of virus injected, as expected. However, in contrast to the pro-resilience effect of PLC SST neuron activation (Fig. 2C), activation of PLC PV neurons exacerbated the CVS effect and led to a reduction in sniffing time in hM3Dq- vs. control vector-injected, CVS-exposed mice (Fig. 2H). Moreover, in contrast to the anxiolytic-like effect of PLC SST neuron activation in the NSFT (Fig. 2C), activation of PLC PV neurons during CVS exposure had anxiogenic-like effects, as evidenced by the additional increase in the latency to feed in hM3Dq- vs. control virus-injected mice (Fig. 2H). Thus, the pro-resilience effect of GABAergic inhibition is specific to dendrite targeting GABAergic inhibition and not observed with PV-neuron mediated inhibition that targets the somatodendritic compartment of pyramidal cells. Notably, the behaviors of hM3Dq-transduced PVCre mice in the EPM (Fig. 2H) and OFT (sFigure 2F) were unaltered, suggesting that the anxiogenic-like effects of PV neuron activation are manifested selectively under acutely stressful conditions.
SST neurons in the PLC and vHPC act sex specifically to reverse chronic stress-induced defects in motivated behavior in male and female mice, respectively.
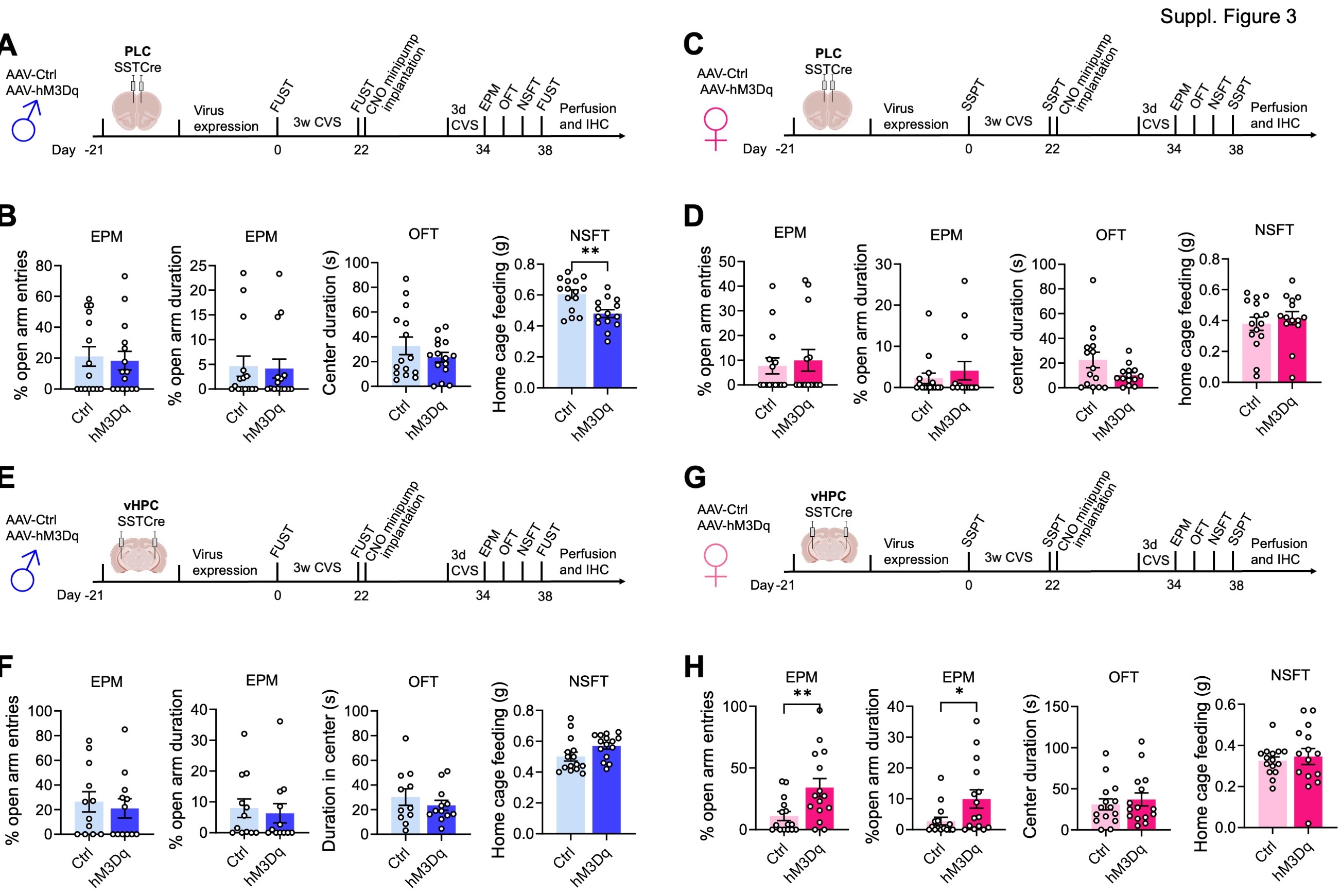
We next asked whether activation of SST neurons in the PLC would reverse prior CVS-induced defects in motivated behavior. Male mice transduced with hM3Dq or control virus were subjected to an initial FUST to assess baseline behavior, followed by three weeks of CVS and a second FUST to verify the stress effects (Fig. 3A). We then implanted CNO minipumps, followed 12 days later with a final behavioral test battery, including a third FUST. To prevent the possible fading of stress effects during the extended time course of experimentation we preceded the final behavioral assessment by three days of CVS exposure, which by itself does not affect motivated behavior 30. Comparison of PLC control- and hM3Dq-vector-injected mice in the FUST before CNO treatment revealed an overall stress effect, independent of the vector injected (Fig. 3B, 1st panel). Comparison of CVS-exposed mice before and after CNO treatment in the second and third FUST revealed an overall CNO effect with a significantly greater Female Urine Sniffing Time in hM3Dq vs. control injected mice (Fig. 3B, 2nd panel). Thus, chronic activation of PLC SST neurons effectively reversed the anhedonia-like consequences of CVS exposure. The behavior in the EPM and OFT remained unaffected, indicating unaltered locomotion and anxiety-related behavior under low-stress conditions (sFigure 3A, B). However, the latency to feed in the NSFT (a comparatively high-stress situation) was significantly reduced in hM3Dq- vs. control-vector-injected mice, indicating a reversal of stress-induced anxiety-like behavior. The home cage feeding of hM3Dq-injected mice was reduced, indicating that the change in the latency to feed was not explained by an altered feeding drive (sFigure 3B, last panel).
Analogous testing of female mice and substitution of SSPTs for the FUSTs and comparing the grooming time before and after CVS exposure revealed a significant stress effect independent of the vector injected (Fig. 3C, D, first panel). However, CNO did not affect the grooming time, independent of the vector injected (Fig. 3D, 2nd panel). Similarly, a pairwise comparison of control and hM3Dq injected mice after CVS and CNO exposure in the NSFT showed no difference between control and hM3Dq injected mice. Therefore, in contrast to male mice, activation of SST neurons in the PLC of female mice failed to reverse CVS-induced anhedonia- and anxiolytic-like behavior. Notably, the behavior in the OFT and EPM was unaltered and consistent with unaltered locomotion (sFigure 3C, D).
The findings thus far suggested that similar to effects on motivated behavior in the absence of stress, the reversal of chronic stress effects by GABAergic inhibition was sex and brain region-specific. To further establish this, we tested the ability of GABAergic circuits in the vHPC to reverse the effects of prior stress exposure (Fig. 3E, F). We injected the vHPC of SSTCre mice with control or hM3Dq virus, exposed the mice to CVS, and then implanted CNO-releasing minipumps (Fig. 3E). Comparison of male mice in the FUST before and after CVS exposure revealed an overall stress effect, independent of virus transduced, as expected (Fig. 3F, first panel). However, in contrast to manipulation of the PLC, CNO exposure of hmD3q injected animals failed to reverse the CVS effect (Fig. 3F, 2nd panel). Similarly, a comparison of control- and hM3Dq-injected mice after CVS and CNO exposure failed to show anxiolysis in the NSFT of hM3Dq-injected mice (Fig. 3F, 3rd panel). By contrast, analogous testing of female mice injected in the vHPC (Fig. 3G) and analyzed in the SSPT revealed a significant CVS effect that was reversed by CNO exposure selectively in hM3Dq- but not control-injected mice (Fig. 3H, 1st and 2nd panel). Moreover, testing in the NSFT revealed an anxiolytic-like effect of CNO selectively in hM3Dq but not control-injected mice (Fig. 3H, 3rd panel). Thus, the reversal of chronic stress-induced anxiety and anhedonia-like behavior in male and female mice mediated by GABAergic microcircuits involves distinct sex-specific cortical substrates.
SST neuron-mediated reversal of CVS effects on motivated behavior involves increased activity of pyramidal cells.
GABAergic synaptic transmission is widely known to inhibit glutamatergic depolarizing inputs and hence to reduce dendritic spiking and output of glutamatergic target neurons 39. By contrast, all evidence suggests that antidepressant drug-like changes in motivated behavior involve increased rather than reduced activity of cortical output neurons 18, 20, 21. We hypothesized, therefore, that chronically and moderately elevated GABAergic transmission at dendrites may lead to increased rather than reduced activity of pyramidal target cells. To test this prediction, we quantitated the densities c-Fos and FosB-expressing neurons as proxies for cell-specific neural activity in vivo. Male mice that had been transduced with AAV-hM3Dq or control virus in the PLC, followed by CVS exposure and CNO-induced reversal of behavioral stress effects (Fig. 3A, 4A), were subjected to immunostaining for c-Fos and virus-encoded, Cre dependent mCherry to assess the density of c-Fos+, mCherry+, as well c-Fos+ non-SST (mCherry-negative) cells representative of pyramidal cells in areas exhibiting virally transduced SST cells. Precisely 90 min before euthanasia the mice were exposed to a novel arena for 10 min to induce a mild but behaviorally relevant arousal state (Fig. 4A). The density of c-Fos+ SST neurons was increased in hM3Dq- compared to control virus-transduced mice, which confirms that chronic low-level CNO-delivery increased the activity of hM3Dq expressing SST neurons (Fig. 4B1,2) with 18.7% and 29.0% of SST neurons showing c-Fos expression in ctrl and hM3Dq injected mice, respectively. Importantly, the density of c-Fos+ non-SST (mCherry-negative) cells was increased in parallel (Fig. 4B3), indicating that chronically increased inhibition by SST cells leads to paradoxically increased activation of non-SST cells. The density of c-Fos+ PV+ cells was less than 5% of the density of c-Fos+ non-SST cells, indicating that c-Fos+ non-SST cells consist predominantly of pyramidal cells. To corroborate these findings, we quantitated the density of SST neurons expressing FosB, a marker that is induced by neuronal depolarization and Ca2+ entry similar to c-Fos but with a slower and longer-lasting time course 40. The density of FosB+ SST cells was increased in hM3Dq vs. control injected mice, with 7.9% and 20.2% of SST neurons showing expression of FosB in control and hM3Dq virus injected mice, respectively (Fig. 4C). Moreover, the density of FosB+ non-SST cells was increased in hM3Dq vs. control-injected mice (Fig. 4C), thereby confirming that SST neurons increase the activity of their putative pyramidal target cells.
Optogenetic stimulation of the large majority of cortical SST neurons results in inhibition of pyramidal cells 41. However, a small subset of SST+ neurons in cortical layer IV has been shown to predominantly inhibit PV+ cells 41, which raised the question of whether the SST neuron-mediated activation of pyramidal cells reported here involved disinhibition via PV cells. We found that the density of c-Fos and PV double-positive neurons was unaltered (Fig. 4D), indicating that SST neuron-mediated activation of pyramidal cells cannot be explained by PV neuron-mediated disinhibition. It follows, that the antidepressant-like, SST neuron-mediated reversal of chronic stress-induced changes in motivated behavior can only be explained by direct SST neuron-mediated activation of pyramidal cells. A proposed mechanism is discussed.
{kind=link}
{kind=link}
{kind=link}