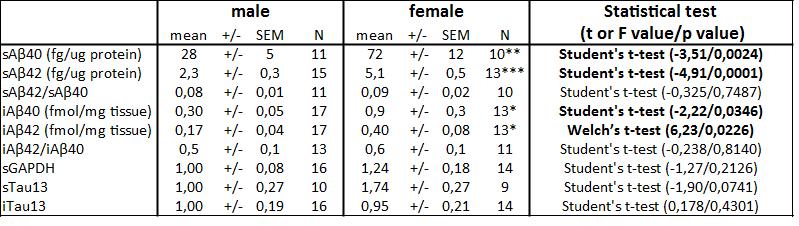
Female 3xTg-AD mice showed more pronounced Aß pathology without significant change in insoluble tau deposits
Studies performed in 3xTg-AD mice have reported differences according to biological sex, particularly for behavior and accumulation of Aß [17–19]. To investigate the basis of this sexual dimorphism, we first quantified molecular factors associated with Aß and tau pathologies in the parietotemporal cortex of 20-month-old heterozygous 3xTg-AD mice. As expected, we observed a more pronounced Aß pathology in both soluble (Aß40: t=-3.51, p=0.0024; Aß42: t=-4.91, p<0.001) and insoluble fractions (Aß40: t=-2.22, p=0.035; Aß42: F=6.23, p<0.023) from female 3xTg-AD mice (Fig. 1). On the other hand, sex difference in the amount of tau in soluble (t=-1.90, p=0.074) and insoluble (t=0.178, p=0.430) fractions did not reach statistical significance (Fig. 2).
Sex-dependent alterations of passive properties by 3xTg-AD expression
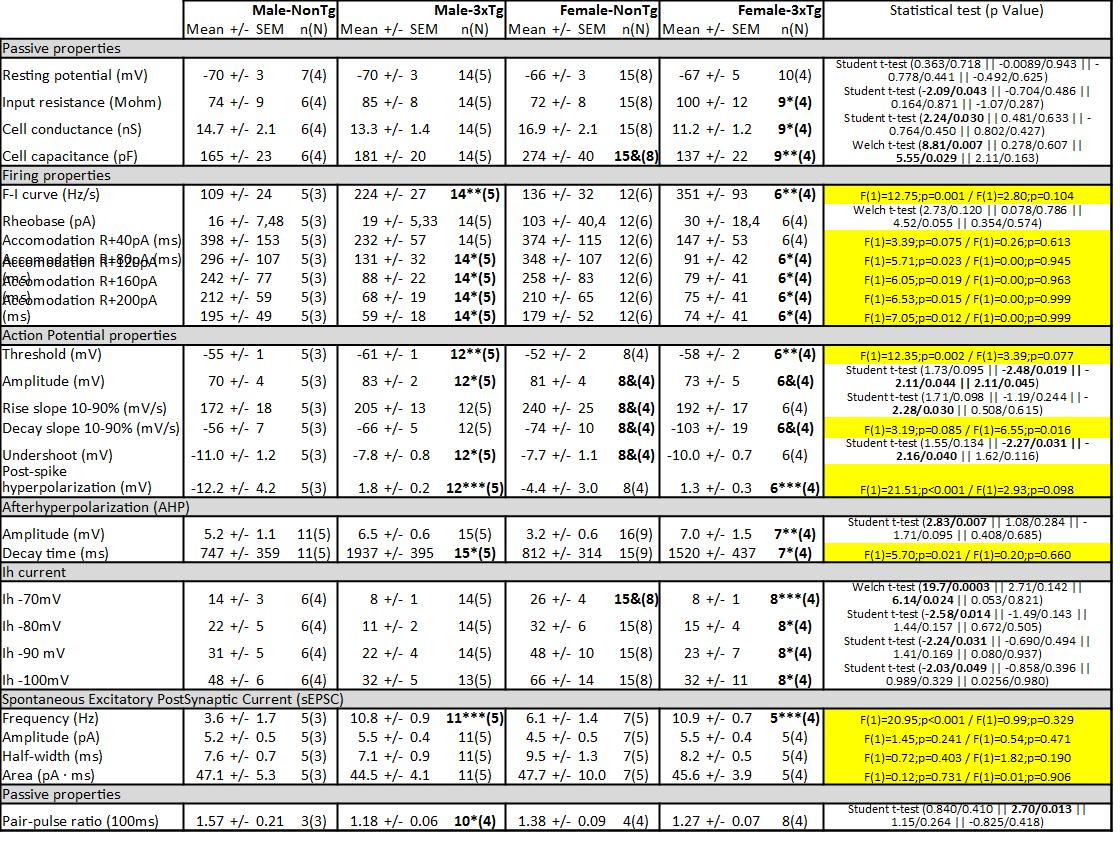
Using a patch-clamp approach (Fig. 3), we investigated the passive properties (Fig. 4A) of layer 5 EC neurons by injecting hyperpolarizing current (Fig. 4B). First, the resting potential was not affected by the sex of animals (Student t-test, NonTg: t=0.778 / p=0.441, 3xTg-AD: t=-0.492 / p=0.625) or by transgene expression (Student t-test, males: t=-0.009 / p=0.943; females: t=0.363 / p=0.718) (Fig. 4C). Second, transgene expression increased the input resistance (Fig. 4D) and decreased the Gc (Fig. 4E) only in females (Student t-test, t=-2.09, p=0.043 for the input resistance; t=2.24, p=0.030 for Gc). Third, we observed a higher CC in NonTg females, compared to female 3xTg-AD (Welch t-test, F=8.81, p=0.007) or to NonTg males (Welch t-test, F=5.55, p=0.029). In addition, the CC correlated with levels of insoluble tau (r2=0.5327, p=0.009, N=23). In accordance with this latter observation, our previous work showed an inverse relationship between the CC and the phosphorylation of tau in 12-month-old homozygous 3xTg-AD mice [35], supporting a link between this electrophysiological property and tau pathology.
Transgene expression was associated with increased firing activity of EC in both sexes
The input-output relationship notifies about how neurons code information in the brain. To investigate this electrophysiological property, we performed steps of depolarizing current (input) and quantified three fundamental features of the firing activity transmitted by neurons: the “Firing rate-Injected current (F-I)” curves, the intensity of depolarization required to deliver an AP (rheobase) and the accommodation (Fig. 5). We found higher F-I curves (two-way ANOVA, genotype: F(1)=12.75 / p=0.001, Fig. 6A) and a lower accommodation (two-way ANOVA, genotype: F(1)=5.71 / p=0.023 for current of +80pA over the maximum current injected in a neuron without reaching its excitation threshold, F(1)=6.05 / p=0.019 for current of +120 pA, F(1)=6.53 / p=0.015 for current of +160 pA, F(1)=7.05 / p=0.012 for current of +200 pA, Fig. 6B) in 3xTg-AD mice of both sexes. The graphically calculated rheobase was not significantly different between each group (Fig. 6C). A positive relationship was observed between F-I curves and insoluble Aß42 (r2=0.5674, p=0.03, N = 19). This transgene-dependent increase of firing activity was in accordance with what we previously reported in 12-month-old 3xTg-AD mice [35].
Sex and transgene expressions were altering AP properties differently
AP is the electrical unit used by neurons to communicate. Our laboratory previously demonstrated that a change in this signal impacts the synaptic response detected by postsynaptic neurons [50], confirming the key role of AP in brain function. In addition, modulation of AP properties is reported during learning processes [51] and aging [37]. To investigate if the conditions used in this study modulated AP properties, we quantified key characteristics associated with a single AP (Fig. 7A, 7B and 7C). Firstly, we observed a decrease in the threshold of APs in 3xTg-AD neurons of both sexes (two-way ANOVA, genotype: F(1)=12.75 / p=0.001, Fig. 7D). Similar results were reported in neocortical layer II/III pyramidal cells from APP transgenic model of AD [52]. Secondly, we found a lower amplitude (Fig. 7E) and a higher undershoot (Fig. 7F) in neurons from 3xTg-AD males compared to NonTg animals of the same sex (Student t-test, t=-2.48 / p=0.019 for amplitude and t=-2.27 / p=0.031 for undershoot). These changes demonstrate a gain in hyperpolarizing currents making up the AP in comparison with depolarizing ones. Thirdly, the rising slope was faster in female NonTg neurons compared to male NonTg cells (Student t-test, t=2.28 / p=0.030, Fig. 7G) and the females of both genotypes showed a higher decay slope than males (two-way ANOVA, genotype: F(1)=6.55 / p=0.016, Fig. 7H). Finally, the post-spike AHP was lower in NonTg neurons versus 3xTg-AD neurons in both males and females (two-way ANOVA, genotype: F(1)=21.51 / p<0.001). We also observed a negative association between the level of insoluble Aß42 and the post-spike AHP (r2 = -0.5174, p = 0.0334, N = 17). Interestingly, these sex-edependent changes in amplitude and undershoot of AP were not reported in homozygous 3xTg-AD mice aged of 12 months [35], showing that specific conditions are necessary to discriminate a difference of sex in these two parameters.
Transgene expression modulated the post-burst AHP differently between males and females
Post-burst AHP potential is known to play a key role in cognitive function by modulating neuronal excitability during learning processes [53–56]. In addition, many studies reported an amplification of this current during aging [54–57] and in AD [58]. To investigate this current, we triggered 2–3 APs by injecting 50 ms depolarizing current and quantified the peak and the decay time of the AHP potential. This current was abolished if calcium was removed from the extracellular solution (Fig. 8A and 8B). We observed an elevation of the amplitude with transgene expression only in females (Student t-test, t=2.83 / p=0.007, Fig. 8C, 8D and 8E), whereas the decay time was longer in 3xTg-AD neurons of both sexes (two-way ANOVA, genotype: F(1)=5.70 / p=0.021, Fig. 8C, 8D and 8F). This current has never been quantified in previous studies using 3xTg-AD mice.
Sex-dependent reduction of Ih current by transgene expression
HCN proteins are subunits known to generate the Ih channel [59]. A previous study reported that HCN1 can form a complex with APP in the murine brain and levels of this protein are significantly reduced in the brains of sporadic AD patients compared with age-matched healthy subjects [60]. In addition, overexpression of HCN1 in Neuro2a cells decreases Aβ generation, whereas blockage of Ih channel activity restores the level of Aβ production [60]. Put together, these data suggest a role of HCN in AD. In this study, we investigated the current generated by HCN channels (Ih) in deep-layer EC neurons from 3xTg-AD mice. The current generated by this channel was quantified by a protocol including steps of hyperpolarized voltage (from -60 mV to -70/-80/-90/- 100 mV) and an application of the antagonist ZD7288 (20 µM) [61] blocked the current (Fig. 9A and 9B). We observed a lower Ih current in female 3xTg-AD neurons when compared to NonTg neurons from the same sex (Welch t-test for step to -70 mV, F=19.7 / p=0.0003, Student t-test for other steps, t=-2.58 / p=0.012 for step to -80 mV, t=-2.24 / p=0.031 for step to -90 mV, t=-2.03 / p=0.049 for step to -100 mV, Fig. 9C to 9E). Moreover, males showed a lower Ih current than females for a voltage step from -60 mV to - 70 mV in NonTg animals (Welch t-test, F=6.14 / p=0.024, Fig. 9E). Finally, the level of sAß42 and iAß40 negatively correlated with the Ih current generated by a step from - 60 mV to - 80 mV (r2 = -0.6368, p = 0.014, N = 14 for sAß40; r2 = -0.4557, p = 0.043, N = 20 for iAß40), supporting the link between Aβ pathology and Ih current.
Basal excitatory synaptic activity was increased by transgene expression in both sexes
Brain hyperactivity and defective network activity were reported in transgenic models of AD neuropathology [34,52,62] and in AD patients [63–65]. We previously quantified the sEPSC of EC neurons and found that these from 12-month-old homozygous 3xTg-AD mice displayed more sEPSC than NonTg without any sex effect [35], which supports the idea of a persistent hyperactivity of glutamatergic synapses in AD. In this study, we reinvestigated the sEPSC in older heterozygous 3xTg-AD mice. Our observations were similar to those earlier obtained, that is an increase in the number of excitatory postsynaptic events by transgene expression with no effect of sex (two-way ANOVA, genotype: F(1)=20.95 / p<0.001, sex: F(1)=0.99 / p=0.329, Fig. 10). A positive relationship was observed between the frequency of sEPSC and iTau (r2 = 0.5392, p = 0.031, N = 16).
Short-term plasticity of cortico-cortical excitatory input was modulated in male 3xTg-AD mice, but not in females
Postsynaptic responses are crucial electrophysiological properties of neurons and depend on how APs are regulated [50,66–68]. When two bursts of AP activate synaptic transmission in a short period, the second postsynaptic response can be larger or smaller than the first. The ratio of the amplitude from the second response to that of the first is called PPR and depends on the probability of vesicular release at the synapse [69]. Then, PPR is used to measure the release probability of cortico-cortical excitatory synapses from fibers of layer 1–2 to dendrites of neurons localized in the layer 5 of EC neurons (Fig. 3C). To discriminate between excitatory and inhibitory inputs, we performed electrical stimulation at different imposed voltages (Fig. 11A and 11B). An excitatory input generated a depolarizing current, which increased upon the application of a hyperpolarized voltage, whereas inhibitory inputs produced a hyperpolarizing current when the imposed voltage was kept under -63 mV (i.e. the estimated reversal potential of Cl− ions) to generate a depolarizing current under that potential. Examples of PPR recorded in neurons of male and female mice expressing or not the transgenes (Fig. 11C). We found a lower PPR in male 3xTg-AD neurons, compared to NonTg cells from the same sex (Student t-test, t=2.70 / p=0.03, Fig. 11E). No difference of genotype was observed in females. Applications of GABAa receptor antagonist picrotoxin (100 µM) with or without NMDA receptor antagonist D-APV (100 µM) in extracellular solution demonstrated that these two receptors were not involved in the postsynaptic current generated in this protocol (Fig. 11F). PPR negatively correlated with sEPSC (r2 = -0.735, p = 0.0005, N = 18) and with F-I curves (r2 = -0.5649, p = 0.022, N = 16), two factors reflecting the neuronal hyperactivity.
Transgenic expression induced synaptic protein impairments
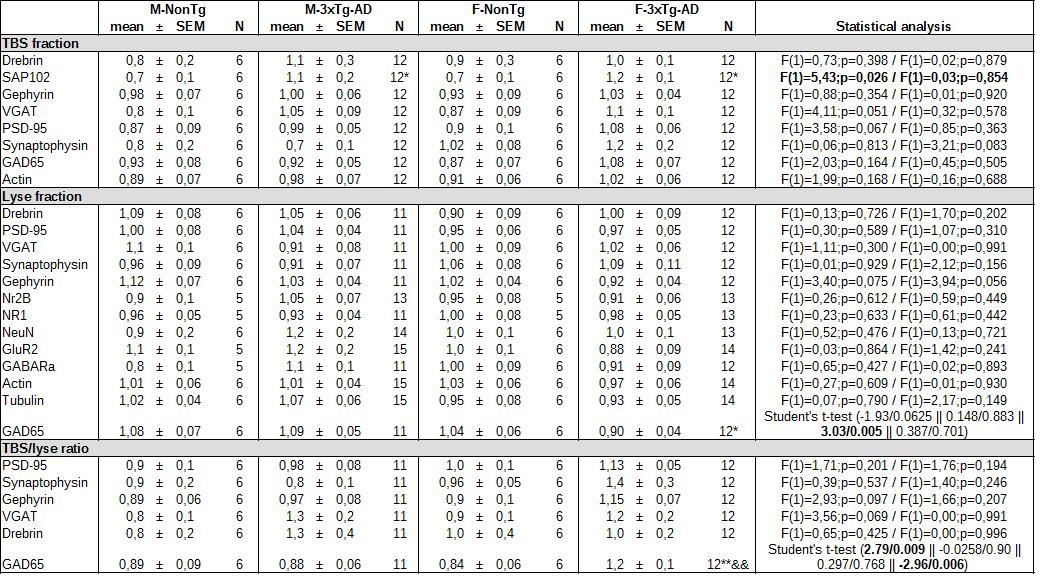
AD is associated with changes in synaptic proteins [35,43,70–75], which could have significant impact at the cellular level. To find if sexes influenced synaptic function at the molecular level in AD, we quantified several proteins in the parietotemporal cortex by western blot. We found an increase of SAP102 in transgenic mice of both sexes (two-way ANOVA, genotype: F(1)=5.43 / p=0.026, Table S3). In addition, the cytosol/membrane ratio of GAD65 is increased only in transgenic females (Student t-test, effect of transgenes in females: t=2.79 / p=0.009, effect of sex in 3xTg-AD: t=-2.96 / p=0.006, Table S3). A positive relationship was observed between SAP102 and the cytosol/membrane ratio of GAD65 (r2 = 0.259, p = 0.002, N = 35). There was no difference for drebrin, gephyrin, VGAT, PSD-95, synaptophysin, Nr2B, NR1, NeuN, GluR2 and GABAaR (Table S3).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}