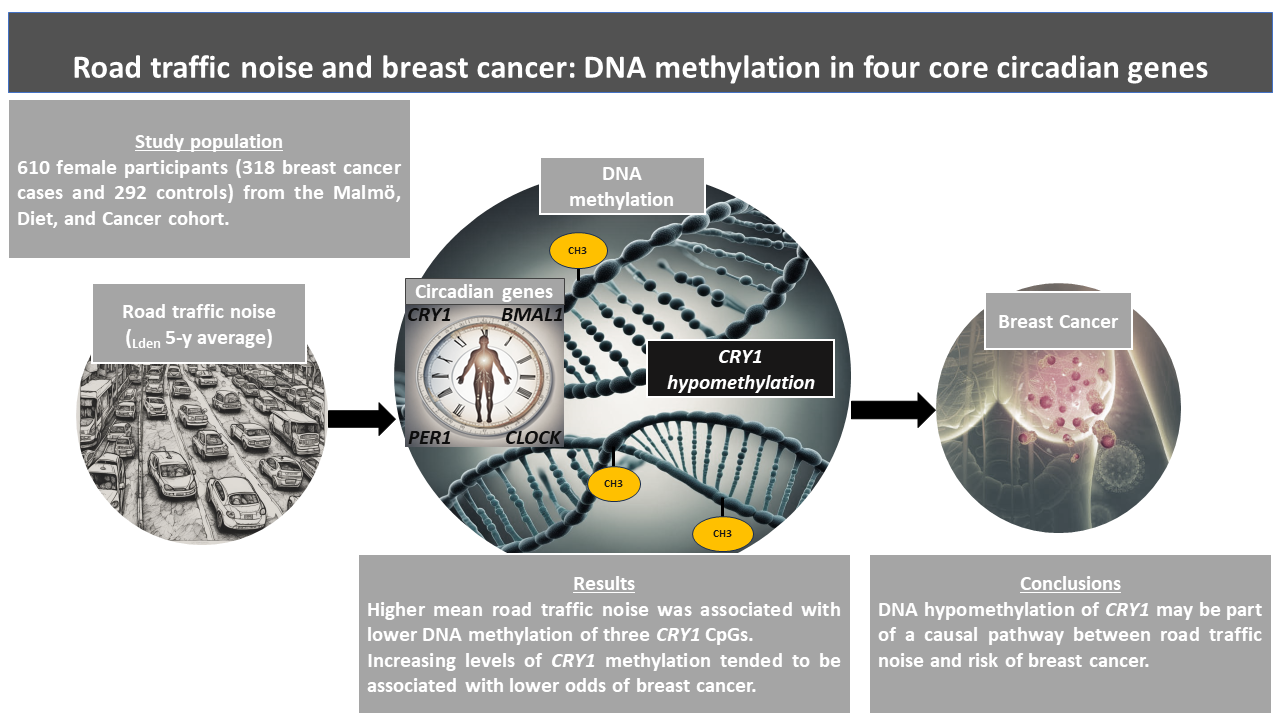
This is the first epidemiological study evaluating the associations between long-term road traffic noise, DNA methylation, and breast cancer. Road traffic noise appeared to be inversely associated with regional changes of CRY1 and BMAL1, and specifically hypomethylation in CRY1 CpG1, CpG2, and CpG12 as well as BMAL1 CpG2, CpG6, and CpG7. In addition, some indication of DNA methylation being inversely associated with breast cancer risk suggests that DNA hypomethylation in certain circadian genes may be part of a causal chain from road traffic noise to breast cancer pathogenesis.
Epidemiological studies examining the association between transportation noise and epigenetic changes are limited. In a Swiss EWAS study, traffic noise demonstrated primarily decreased methylation at specific DMRs, which is somewhat in line with the present study where we found road traffic noise to be associated with hypomethylation in multiple CRY1 and BMAL1 CpGs (40). Additionally, in the brains of rats, long-term nocturnal noise was associated with aberrant methylation, in particularly hypomethylation of the melanocortin 2 receptor (Mc2r) gene in the hippocampus (41). Further evidence from murine models demonstrated that murine cochlea and inferior colliculus contain circadian machinery, and that noise exposure differentially impacted the expression of core clock genes in the auditory periphery and inferior colliculus (42, 43). Both CRY1 and BMAL1 are core components of the circadian clock, along with other period genes, and orchestrate the circadian rhythm through the complex interplay involving positive and negative feedback loops, self-expression regulation, as well as additional axillary regulatory processes (44). In short, BMAL and CLOCK transcription factors form the heterodimer that promotes the expression of CLOCK and CLOCK-regulated genes. Conversely, PER and CRY constitute the inhibitory complex which impedes the CLOCK-BMAL protein complex (28, 37).
The consequences of our results need to be elucidated since DNA hypermethylation is frequently linked with transcriptional gene repression, while hypomethylation is often linked with a chromatin arrangement that supports transcription (45). It is unclear what the methylation changes observed in the present study are predicted to result in, but overexpression and aberrant expression of certain circadian genes have been found in cancer tissue, including breast cancer (46). Thus, it is conceivable that long-term road traffic noise could lead to altered gene transcription and expression, hallmarks in multiple cancers, including breast cancer.
We observed some indication that methylation of multiple CpGs in CRY1 and CLOCK were inversely associated with breast cancer. CRY1’s role in breast cancer development is not fully understood, however, CRY is involved in regulation of DNA replication, DNA damage, and cell cycle (47, 48). CRY1 in particular, is also a known regulator of cell proliferation and DNA repair (49), and has been shown to inhibit nuclear receptors involved in certain cancers (50). Two studies demonstrated a link between hypermethylation of the CLOCK gene with lower breast cancer risk which is congruent with our findings (51, 52). Increased methylation might result in reduced gene expression, consequently weakening CLOCK proliferation. Furthermore, CLOCK and CRY1 might possess tumorigenic characteristics, and this is substantiated by whole genome expression microarray studies, that found expression of multiple cancer related transcripts to be modified after CLOCK gene knockdown. More specifically, after silencing the CLOCK gene, the genes primarily involved in breast cancer progression included CCL5 (53), SP100 (54), and BDKRB2 (55).
In summary, the aforementioned factors reveal a potential pathway from road traffic noise to dysregulation of the circadian clock and breast carcinogenesis. Nevertheless, the mechanism from noise to circadian rhythm disruption and the development of breast cancer remains to be fully elucidated.
Road traffic noise and traffic related air pollution are correlated since they share the same emission sources, and air pollution has also been linked to both DNA methylation as well as breast cancer risk (56, 57). Therefore, it is crucial for research on noise exposure to take into consideration air pollution, and conversely, for studies on air pollution to consider traffic noise exposure. In the present study, estimates for road traffic noise and methylation, as well as for DNA methylation and breast cancer, were not impacted to any large extent when adjusting for PM2.5 or NOx.
We opted to not adjust for multiple comparisons since the CpGs are intercorrelated, particularly for CRY1, and therefore would result in overadjustment.
A key strength of the present study is that it is based on a well-characterized cohort which includes data on many potential confounders, namely, air pollution and inconvenient working hours. Another strength is that we focused on DNA methylation in specific genes related to both sleep disturbance and breast cancer. Lastly, we utilized pyrosequencing (DNA sequencing) and is considered the benchmark for analyzing DNA methylation.
Although our findings suggest that long-term road traffic noise potentially results in epigenetic changes in circadian genes, the molecular pathomechanisms underlying this phenomenon remain obscure. An important limitation is that our findings are based on a limited sample size and further studies to corroborate our findings are recommended. Another limitation of our study is that we measured DNA methylation in lymphocytes and not the brain or breast. However, circadian clocks are present in most cells throughout the body. Lastly, we lack information on artificial light at night, which could potentially bias our findings, as light at night is associated with disruption of the circadian rhythm and has been purported as a possible mechanism of cancer etiology (58).
{kind=link}