Changes in phyllosphere and rhizosphere microbial community alpha diversity
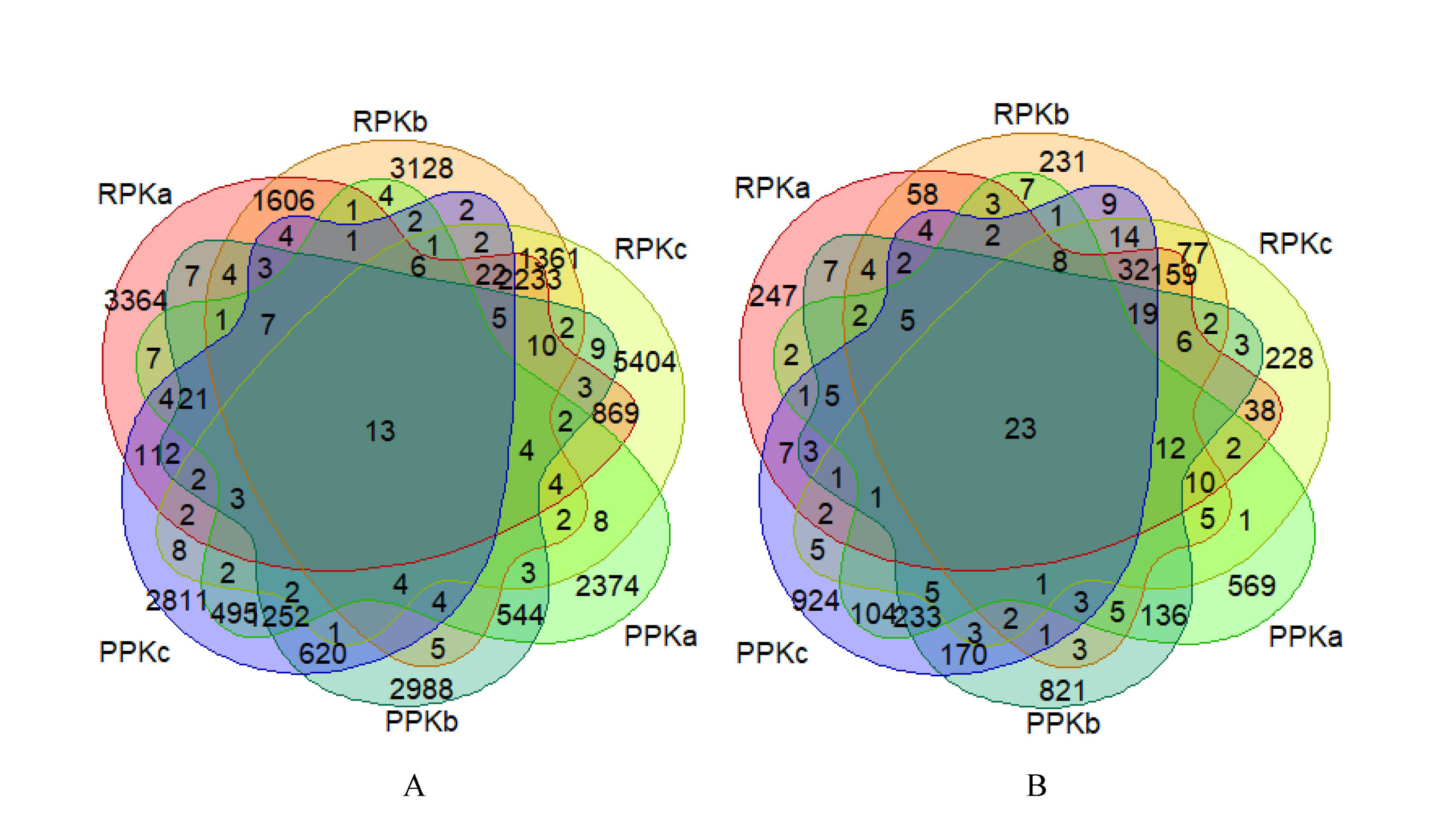
A total of 1,367,262 and 1,174,114 high-quality phyllosphere and rhizosphere bacterial sequences were generated across all samples after sequence denoising and quality filtering with the average number of sequences per sample 91, 150 and 78, 274, severally, which were assigned into 11, 294 and 18, 175 OTUs. The number of shared phyllosphere bacterial OTUs among PPKa, PPKb, and PPKc was 1306, and the unique OTUs of PPKa, PPKb, and PPKc was 2,400, 3,028, and 2,862, respectively (Fig. 1A). The number of shared rhizosphere bacterial OTUs among RPKa, RPKb, and RPKc of was 2,297, and the unique OTUs of RPKa, RPKb, and RPKc was 3,416, 3,145, and 5,437, respectively (Fig. 1B). Moreover, the shared OTUs among RPKa, RPKb, RPKc, PPKa, PPKb, and PPKc was 13 (Fig. S2A).
The fungal communities were further explored by high-throughput amplicon sequencing. Across all samples, we obtained a total of 1, 318, 977 and 1, 316, 090 high-quality phyllosphere and rhizosphere fungal sequences after sequence denoising and quality filtering with the average number of sequences per sample 87, 739 and 87, 931, severally, which were respectively grouped into 1, 272 and 3, 190 OTUs. The number of shared phyllosphere fungal OTUs among PPKa, PPKb, and PPKc was 276, and the number of unique OTUs of PPKa, PPKb, and PPKc was 599, 846, and 997, respectively (Fig. 1C). The number of shared rhizosphere fungal OTUs among RPKa, RPKb, and RPKc was 269, and the number of unique OTUs of RPKa, RPKb, and RPKc was 272, 260, and 245, severally (Fig. 1D). In addition, the shared OTUs among RPKa, RPKb, RPKc, PPKa, PPKb, and PPKc was 23 (Fig. S2B).
As expected, there was considerable variation of phyllosphere bacterial Pielou_e (F= 12.639, P= 0.001), Shannon (F= 10.268, P= 0.003), and Simpson index (F = 5.882, P = 0.017) among PPKa, PPKb, and PPKc. Furthermore, relative to PPKa and PPKb, PPKc increased phyllosphere bacterial Pielou_e, Shannon, and Simpson index with 0.72, 7.83, and 0.97, respectively (Table 1). In addition to Goods_coverage (F = 3.533, P = 0.062) and Simpson index (F = 3.235, P = 0.075), rhizosphere bacterial Observed_species (F = 6.777, P = 0.011), Chao 1 (F = 4.655, P = 0.032), Pielou_e (F=47.496, P=0.0001), and Shannon index (F = 11.772, P = 0.002) were observed significant differences among RPKa, RPKb, and RPKc. What’s more, RPKc hold the highest rhizosphere bacterial Observed_species, Chao 1, Pielou_e, Shannon, and Simpson index with 5384.50, 6916.43, 0.895, 11.09, 0.9987, separately (Table 2). With regard to fungi, phyllosphere fungal Chao 1 index (F= 56.306, P= 0.000), Goods_coverage (F= 14.509, P= 0.001), Observed_species (F= 56.689, P= 0.000) differed dramatically among PPKa, PPKb, and PPKc, and PPKc hold the highest Chao1 index with 597.46 (Table 1). Rhizosphere fungal Pielou_e (F= 12.639, P= 0.001), Shannon (F= 10.268, P= 0.003), and Simpson index (F= 5.882, P= 0.017) among RPKa, RPKb, RPKc also appeared obviously different. It is well established that RPKc owned highest rhizosphere fungal Pielou_e, Shannon, and Simpson index with 0.56, 4.66, and 0.89 (Table 2).
Variations in phyllosphere and rhizosphere microbial community beta diversity
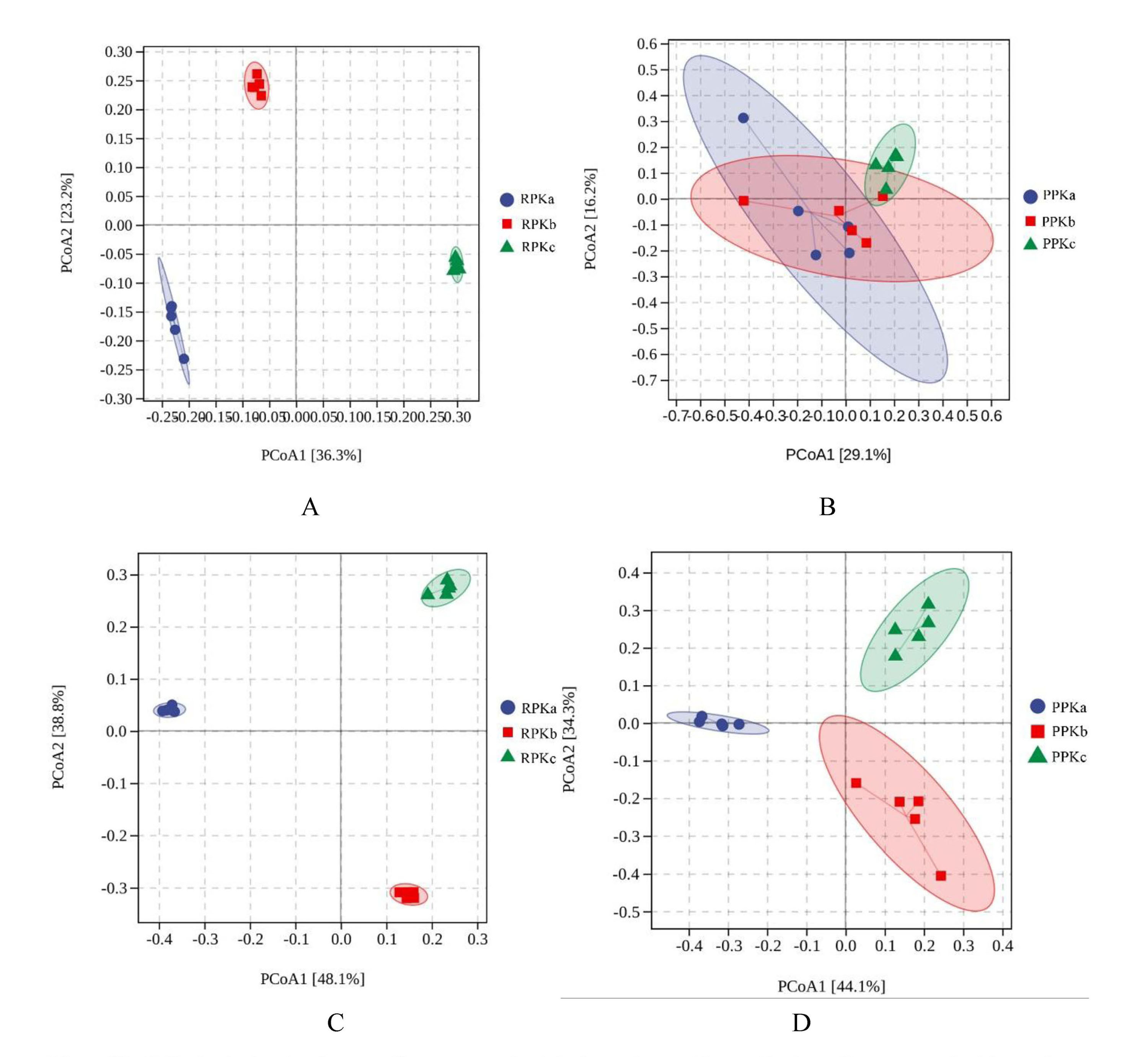
It is well established that the microbial compositions from rhizosphere and phyllosphere samples formed distinct clusters (Fig. 2), which indicated that the plant compartment is a major selective force for the formation of plant-related microbial composition. The unconstrained principle coordinate analysis (PCoA) of bray-curits distance from all phyllosphere and rhizosphere samples based on the OTU data detected 64.9% of the total variance among bacterial communities, with the first and second axes explaining 57.2% and 7.7% of the variance, respectively (Fig. 2A). PCoA analysis based on the OTU data detected 67.0% of the total variance of fungal communities, with the first and second axes explaining 55.7% and 11.3% of the variance, respectively (Fig. 2B). As expected, the infection of B. xylophilus had a profound effect on plant microbe. The results demonstrated that rhizosphere bacterial community (Fig. S3A), rhizosphere fungal community (Fig. S3C), and phyllosphere fungal community (Fig. S3D) from PKa, PKb, and PKc formed three distinct clusters, especially along the PCoA1.
Comparative analysis of phyllosphere and rhizosphere microbial community composition
For bacteria, at the phylum level, 36 rhizosphere bacterial groups were obtained, and 8 bacterial communities with the relative abundance more than 1% were detected, including Proteobacteria, Actinobacteria, Acidobacteria, Verrucomicrobia, Chloroflexi, Gemmatimonadetes, Patescibacteria, Bacteroidetes, and Firmicutes, accounting for 94.75% (Fig. 3A). 27 phyllosphere bacterial groups were obtained, and 4 bacterial communities with the relative abundance more than 1% were obtained, including Proteobacteria, Actinobacteria, Bacteroidetes, and Cyanobacteria (Fig. 3B). As for fungi, at the phylum level, 7 rhizosphere fugal groups were obtained, and 2 fungal communities with the relative abundance more than 1% were detected, including Ascomycota and Basidiomycota (Fig. 4A). 14 phyllosphere fungal groups were obtained, and 4 fungal communities with the relative abundance more than 1% were detected, including Basidiomycota, Ascomycota, Mortierellomycota, and Mucoromycota (Fig. 4B).
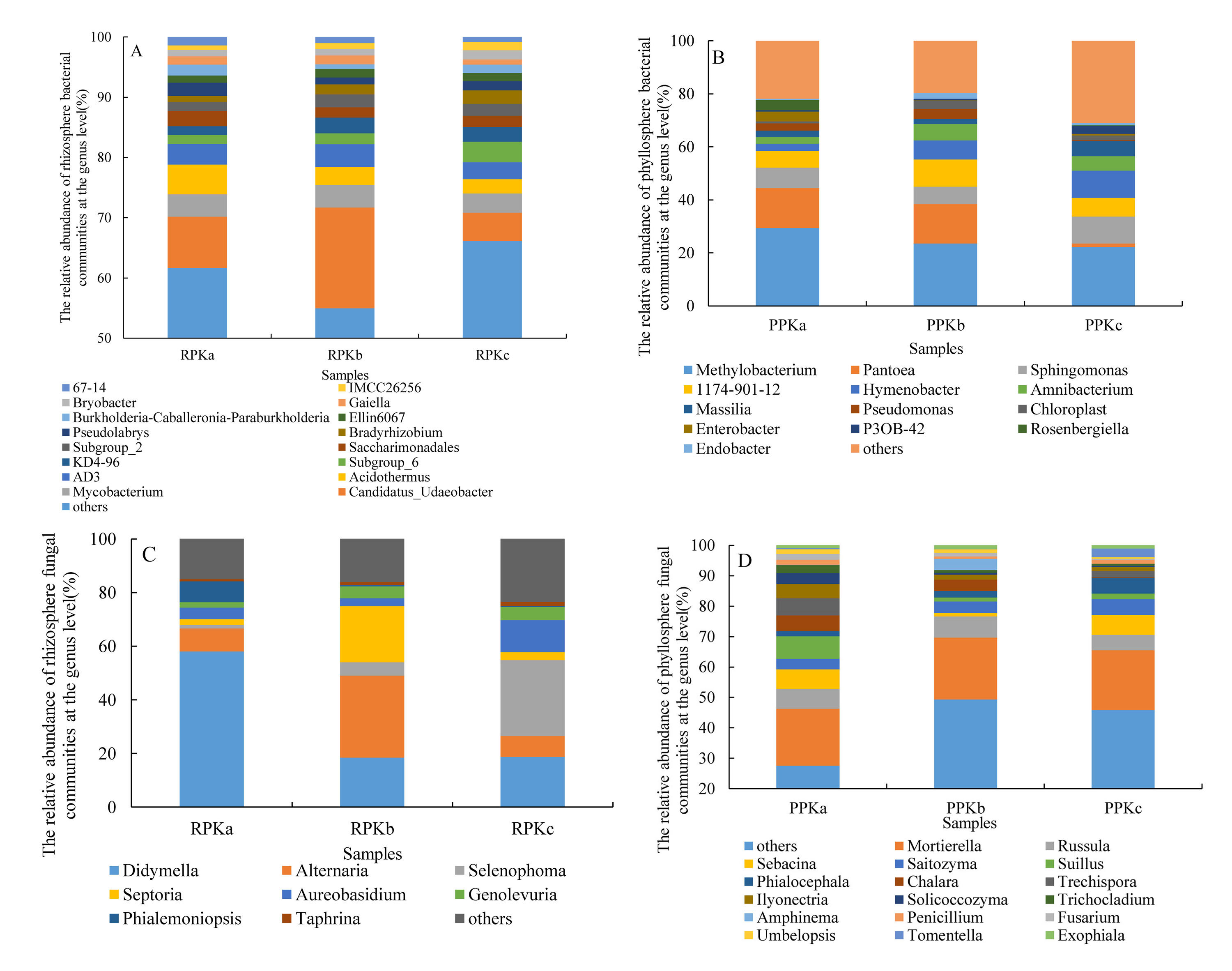
At the genus level, 851 rhizosphere bacterial communities were obtained, of which, the average relative abundance of Candidatus_Udaeobacter, Mycobacterium, Acidothermus, AD3, Subgroup_6, KD4-96, Saccharimonadales, Subgroup_2, Bradyrhizobium, Pseudolabrys, Ellin6067, Burkholderia-Caballeronia-Paraburkholderia, Gaiella, Bryobacter, IMCC26256, and 67-14 was more than 1% (Fig. S4A). 606 phyllosphere bacterial community were obtained, and the relative abundance of Methylobacterium, Pantoea, Sphingomonas, 1174-901-12, Hymenobacter, Amnibacterium, Massilia, Pseudomonas, Chloroplast, Enterobacter, P3OB-42, Rosenbergiella, and Endobacter was more than 1% (Fig. S4B). Heatmap demonstrated that rhizosphere (Fig. 5A) and phyllosphere (Fig. 5B) bacteria from RPKa (PPKa) and RPKb (PPKb) formed a cluster, clearly distinguished from those of RPKc (PPKc). For fungi, 321 rhizosphere fungal communities were obtained, among which, the groups with the relative abundance more than 1% were Didymella, Alternaria, Selenophoma, Septoria, Aureobasidium, Genolevuria, Phialemoniopsis, and Taphrina (Fig. S4C). 492 phyllosphere fungal community were obtained, and the groups with the relative abundance more than 1% were Mortierella, Russula, Sebacina, Saitozyma, Suillus, Phialocephala, Chalara, Trechispora, Ilyonectria, Solicoccozyma, Trichocladium, Amphinema, Penicillium, Fusarium, Umbelopsis, Tomentella, and Exophiala (Fig. S4D). Heatmap demonstrated that rhizosphere (Fig. 5C) and phyllosphere (Fig. 5D) fungi from RPKb (PPKb) and RPKc (PPKc) formed a cluster, clear distinguished from those of RPKa (PPKa).
Furthermore, we conducted LEfSe analysis to identify which microbial taxa (from phylum to genus level) were major contributors to the differences in rhizosphere and phyllosphere community compositions among different samples (Fig. 6). At the phylum level, the larger groups of rhizosphere bacteria in RPKa were Actinobacteria, Gemmatimonadetes, and Patescibacteria, while in RPKb were Chloroflexi, Rokubacteria, and Verrucomicrobia, in RPKc were Acidobacteria, Bacteroidetes, and Proteobacteria (Krus-kall-Wallis test, P < 0.05) (Fig. 6A). The larger group of phyllosphere bacterial group in PPKa was Proteobacteria, while in PPKb were Acidobacteria, Chloroflexi, and Cyanobacteria, in PPKc were Actinobacteria, Armatimonadetes, Bacteroidetes, and Planctomycetes (P < 0.05) (Fig. 6B). Additionally, as for fungi, the RPKa contained a significantly higher abundance of Phialemoniopsis than RPKb and RPKc samples (P < 0.05). While, RPKb owned higher abundances of Ascomycota and Pseudovirgaria. Curvibasidium, Neophaeococcomyces, Selenophoma, and Symmetrosporaceae presented higher in RPKc (Fig. 6C). For phyllosphere, the fungi group of Chalara, Basidiomycota, Mucoromycota, and Staphylotrichum were significantly enriched in PPKa, while Rozellomycota, and Arthrocatena were more enriched in PPKb as compared to PPKa and PPKc (P < 0.05). The phyla Ascomycota was more abundant in PPKc than PPKa and PPKb (P < 0.05) (Fig. 6D).
Microbiological information network and co-occurrence analysis
In order to a deeper comparative look into the community structure of the microbial taxa, we created association networks of phyllosphere and rhizosphere bacterial and fungal communities from OTU data (Fig.7; Table S1). Total nodes of phyllosphere and rhizosphere bacterial community association network in PKc existed the highest, followed by PKb and PKa, and total nodes of phyllosphere and rhizosphere fungal community association network in PKb existed the highest, followed by PKc and PKa, both indicating that the OTUs of the ecological network increased after infection (Fig.7; Table S1). Graph density in the network of PKb, a key topological property to describe how well a node is connected with its neighbours, showed higher than PKa and PKc, suggestive of more intensive microbial coupling at the early stage of infection (Fig. 7; Table S1). Except for phyllosphere fungi, positive links showed decreased with the infection of PWD, and at the last stage, positive links existed the lowest (Fig. 7; Table S2), demonstrating that most of the microbial taxa tended to be co-excluding rather than co-occurring.
{kind=link}
{kind=link}
{kind=link}
{kind=link}